Вторичная структура ДНК. Правила Чаргаффа.
Изучая состав ДНК, Чаргафф (1949г.) установил важные закономерности соотношения АО в ДНК, что в дальнейшем помогло раскрыть их структуру.
1) сумма пуриновых оснований = сумме пиримидиновых оснований.
А + Г = Ц + Т или 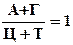
2) соединение А = соединению Т
А = Т или 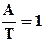
3) соединение Г = соединению Ц
Г = Ц или 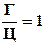
4) сумма А и Ц = сумме Г и Т
А+Ц = Г+Т или 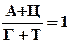
5) изменчива только сумма А+Т и Г+Ц
если А+Т > Г+Ц, то это АТ-тип ДНК
если наоборот, то это ГЦ-тип ДНК.
Эта разница выражается через коэффициент специфичности, который является важной характеристикой молекулы ДНК.
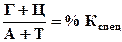 (в % от суммы Г+Ц от 0,54 до 0,94 для живых, то есть преобладает АТ-тип; от 0,45 до 2,57 – для микроорганизмов).
(в % от суммы Г+Ц от 0,54 до 0,94 для живых, то есть преобладает АТ-тип; от 0,45 до 2,57 – для микроорганизмов).
Правила Чаргаффа характерны только для ДНК, но не свойственны РНК.
На основании этих правил и большого числа разнообразных экспериментальных данных в 1953 г. Д. Уотсон и Ф. Крик разработали модель ВС ДНК.
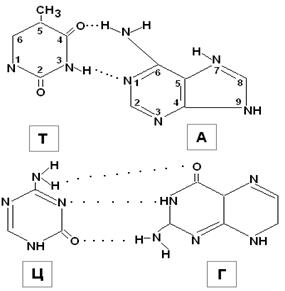
1. Молекула ДНК построена из двух полинуклеотидных. цепей, ориентированных антипараллельно (3′ конец одной цепи соседствует с 5′ концом второй цепи); цепи на всем протяжении связаны Н-связями. При этом каждый мононуклеотид участвует в образовании Н-связей.
2. Н-связи образованы между А одной цепи и Т другой, между Г одной цепи и Ц другой. Основания, образующие такие пары комплементарны друг другу, то есть между ними легче образуется Н-связь.
|
3. ПС одной цепи ДНК комплементарна ПС второй цепи, что не означает их идентичности 
4. Обе цепи закручены вправо в спираль, имеющую общую ось. Цепи могут быть разделены только раскручиванием. Такие спирали называются плектонемическими.
5. Пурины и пиримидиновые основания обращены внутрь спирали. Их плоскости перпендикулярны оси спирали и параллельны друг другу, таким образом получается стопка оснований. Пентозофосфатные остовы обеих цепей обращены наружу от спирали.
Стабилизация ВС ДНК обеспечена за счет:
1) особого вида вандервальсовых сил между АЩ – так называемых стэкинг-взаимодействий;
2) водного окружения фосфатов и особенно нейтрализации их зарядов. –q стремятся оттолкнуться друг от друга, вытягивают цепь ДНК и тем самым стабилизируют ее;
3) образование двойной спирали ДНК возможно только при условии связывания фосфатов (–q) с гистонами – основными белками (+q);
4) Н-связями.
Комплекс белков с ядерной ДНК клеток называется хроматином.
Двойная спираль (b-форма) Уотсона и Крика имеет размерность:
1) a-спирали – 2,1 нм;
2) один виток (шаг спирали) составляет 10 пар АО; 3,4 нм;
3) высота каждого мононуклеотида 0,34 нм.
В природной ДНК хромосом строгая двуспиральность нарушается на тех участках, которые называются палиндромами (перевертышами). В таких фрагментах ДНК последовательность мононулеотидов одинакова слева направо и справа налево. Комплементарные основания палиндромов спариваются и образуют «кресты» или «шпильки». Они помогают регуляторным белкам узнавать места списывания генетического текста ДНК хромосом.
Третичная структура ДНК образуется в результате дополнительного скручивания в пространстве в виде суперспирали или изогнутой, то есть сломанной двойной спирали.
В составе хроматина ДНК сост 30-45%; гистоны 30-50%; негистиновый белок 4-33%; РНК 1,5-10%.
В электронном микроскопе хроматин напоминает бусы – шаровидные утолщения – нуклеосомы, с перемычками. Каждая нуклеосома содержит 8 молекул гистонов и намотанный на них участок ДНК, содержащий 146 пар АО. При такой укладке длина молекулы ДНК уменьшается ~ в 7 раз. Это лишь первый уровень укладки цепи. Длина молекул ДНК 3-5 см, а длина хромосом всего несколько мкм, то есть степень укорочения ДНК достигает нескольких тысяч. Это происходит в результате дополнительного скручивания нуклеосомной нитки бус.
Дата добавления: 2015-12-22; просмотров: 1416;