Глава 3. ВАЖНЕЙШИЕ АБИОТИЧЕСКИЕ ФАКТОРЫ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ 4 страница
3.3.2. Экологические группы растений по отношению к воде
Гидатофиты – это водные растения, целиком или почти целиком погруженные в воду. Среди них – цветковые, которые вторично перешли к водному образу жизни (элодея, рдесты, водяные лютики, валлиснерия, уруть и др.). Вынутые из воды, эти растения быстро высыхают и погибают. У них редуцированы устьица и нет кутикулы. Транспирация у таких растений отсутствует, а вода выделяется через особые клетки – гидатоды.
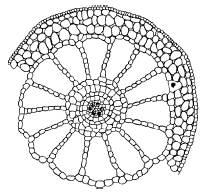
Рис. 32. Поперечный срез стебля урути Myriophyllum verticillatum (по Т. К. Горышиной, 1979)
Листовые пластинки у гидатофитов, как правило, тонкие, без дифференцировки мезофилла, часто рассеченные, что способствует более полному использованию ослабленного в воде солнечного света и усвоению СО2. Нередко выражена разнолистность – гетерофиллия; у многих видов есть плавающие листья, имеющие световую структуру. Поддерживаемые водой побеги часто не имеют механических тканей, в них хорошо развита аэренхима (рис. 32).
Корневая система цветковых гидатофитов сильно редуцирована, иногда отсутствует совсем или утратила свои основные функции (у рясок). Поглощение воды и минеральных солей происходит всей поверхностью тела. Цветоносные побеги, как правило, выносят цветки над водой (реже опыление совершается в воде), а после опыления побеги снова могут погружаться, и созревание плодов происходит под водой (валлиснерия, элодея, рдесты и др.).
Гидрофиты – это растения наземно-водные, частично погруженные в воду, растущие по берегам водоемов, на мелководьях, на болотах. Встречаются в районах с самыми разными климатическими условиями. К ним можно отнести тростник обыкновенный, частуху подорожниковую, вахту трехлистную, калужницу болотную и другие виды. У них лучше, чем у гидатофитов, развиты проводящие и механические ткани. Хорошо выражена аэренхима. В аридных районах при сильной инсоляции их листья имеют световую структуру. У гидрофитов есть эпидерма с устьицами, интенсивность транспирации очень высока, и они могут расти только при постоянном интенсивном поглощении воды.
Гигрофиты – наземные растения, живущие в условиях повышенной влажности воздуха и часто на влажных почвах. Среди них различают теневые и световые. Теневые гигрофиты – это растения нижних ярусов сырых лесов в разных климатических зонах (недотрога, цирцея альпийская, бодяк огородный, многие тропические травы и т. п.). Из-за высокой влажности воздуха у них может быть затруднена транспирация, поэтому для улучшения водного обмена на листьях развиваются гидатоды, или водяные устьица, выделяющие капельно-жидкую воду. Листья часто тонкие, с теневой структурой, со слабо развитой кутикулой, содержат много свободной и малосвязанной воды. Обводненность тканей достигает 80 % и более. При наступлении даже непродолжительной и несильной засухи в тканях создается отрицательный водный баланс, растения завядают и могут погибнуть.
К световым гигрофитам относятся виды открытых местообитаний, растущие на постоянно влажных почвах и во влажном воздухе (папирус, рис, сердечники, подмаренник болотный, росянка и др.). Переходные группы – мезогигрофиты и гигромезофиты.
Мезофиты могут переносить непродолжительную и не очень сильную засуху. Это растения, произрастающие при среднем увлажнении, умеренно теплом режиме и достаточно хорошей обеспеченности минеральным питанием. К мезофитам можно отнести вечнозеленые деревья верхних ярусов тропических лесов, листопадные деревья саванн, древесные породы влажных вечнозеленых субтропических лесов, летнезеленые лиственные породы лесов умеренного пояса, кустарники подлеска, травянистые растения дубравного широкотравья, растения заливных и не слишком сухих суходольных лугов, пустынные эфемеры и эфемероиды, многие сорные и большинство культурных растений. Из приведенного перечня видно, что группа мезофитов очень обширна и неоднородна. По способности регулировать свой водный обмен одни приближаются к гигрофитам (мезогигрофиты), другие – к засухоустойчивым формам (мезоксерофиты).
Ксерофиты растут в местах с недостаточным увлажнением и имеют приспособления, позволяющие добывать воду при ее недостатке, ограничивать испарение воды или запасать ее на время засухи. Ксерофиты лучше, чем все другие растения, способны регулировать водный обмен, поэтому и во время продолжительной засухи остаются в активном состоянии. Это растения пустынь, степей, жестколистных вечнозеленых лесов и кустарниковых зарослей, песчаных дюн.
Ксерофиты подразделяются на два основных типа: суккуленты и склерофиты.
Суккуленты– сочные растения с сильно развитой водозапасающей паренхимой в разных органах. Стеблевые суккуленты – кактусы, стапелии, кактусовидные молочаи; листовые суккуленты – алоэ, агавы, мезембриантемумы, молодило, очитки; корневые суккуленты – аспарагус. В пустынях Центральной Америки и Южной Африки суккуленты могут определять облик ландшафта (рис. 33).

Рис. 33. Суккулентное растение – древовидный кактус из пустыни Аризона
Листья, а в случае их редукции стебли суккулентов имеют толстую кутикулу, часто мощный восковой налет или густое опушение. Устьица погруженные, открываются в щель, где задерживаются водяные пары.
Днем они закрыты. Это помогает суккулентам сберегать накопленную влагу, но зато ухудшает газообмен, затрудняет поступление СО2 внутрь растения. Поэтому многие суккуленты из семейств лилейных, бромелиевых, кактусовых, толстянковых ночью при открытых устьицах поглощают СО2, который только на следующий день перерабатывают в процессе фотосинтеза. Поглощенный СО2 переводится в малат. Кроме того, при дыхании ночью углеводы разлагаются не до углекислого газа, а до органических кислот, которые отводятся в клеточный сок. Днем на свету малат и другие органические кислоты расщепляются с выделением СО2, который используется в процессе фотосинтеза. Таким образом, крупные вакуоли с клеточным соком запасают не только воду, но и СО2. Так как у суккулентов ночная фиксация углекислоты и переработка ее днем в ходе фотосинтеза разделены во времени, они обеспечивают себя углеродом, не подвергаясь риску чрезмерной потери воды, но масштабы поступления углекислого газа при таком способе невелики, и растут суккуленты медленно.
Осмотическое давление клеточного сока суккулентов мало – всего 3 · 105 – 8 · 105 Па (3–8 атм), они развивают небольшую сосущую силу и способны всасывать воду лишь атмосферных осадков, просочившихся в верхний слой почвы. Корневая система их неглубокая, но сильно распростертая, что особенно характерно для кактусов.
Склерофиты– это растения, наоборот, сухие на вид, часто с узкими и мелкими листьями, иногда свернутыми в трубочку. Листья могут быть также рассеченными, покрытыми волосками или восковым налетом. Хорошо развита склеренхима, поэтому растения без вредных последствий могут терять до 25 % влаги не завядая. В клетках преобладает связанная вода. Сосущая сила корней до нескольких десятков атмосфер, что позволяет успешно добывать воду из почвы. При недостатке воды резко снижают транспирацию. Склерофиты можно подразделить на две группы: эуксерофитов и стипаксерофитов.
К эуксерофитам относятся многие степные растения с розеточными и полурозеточными, сильно опушенными побегами, полукустарнички, некоторые злаки, полынь холодная, эдельвейс эдельвейсовидный и др. Наибольшую биомассу эти растения создают в период, благоприятный для вегетации, а в жару уровень обменных процессов у них очень низок.
Стипаксерофиты– это группа узколистных дерновинных злаков (ковыли, тонконоги, типчак и др.). Характеризуются низкой транспирацией в засушливый период и могут переносить особенно сильное обезвоживание тканей. Свернутые в трубочку листья имеют внутри влажную камеру. Транспирация идет через погруженные в бороздки устьица внутрь этой камеры, что снижает потери влаги (рис. 34).
А, Б
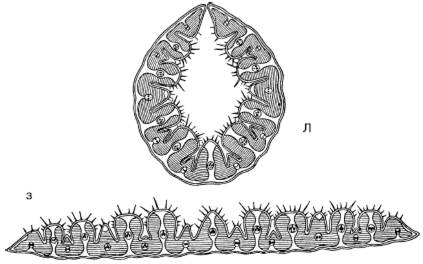
Рис. 34. Поперечный срез листа ковыля Stipa capillata (по A. Кернеру, 1896): A – при засухе (лист свернут); Б – во влажную погоду (пластинка листа развернута)
Кроме названных экологических групп растений, выделяют еще целый ряд смешанных или промежуточных типов.
Различные пути регуляции водообмена позволили растениям заселить самые различные по экологическим условиям участки суши. Многообразие приспособлений лежит, таким образом, в основе распространения растений по поверхности земли, где дефицит влаги является одной из главных проблем экологических адаптаций.
3.3.3. Водный баланс наземных животных
Животные получают воду тремя основными путями: через питье, вместе с сочной пищей и в результате метаболизма, т. е. за счет окисления и расщепления органических веществ – жиров, белков и углеводов.
Некоторые животные могут впитывать воду через покровы из влажного субстрата или воздуха, например личинки некоторых насекомых – мучного хрущака, жуков-щелкунов и др.
Потери воды у животных происходят через испарение покровами или со слизистых оболочек дыхательных путей, путем выведения из тела мочи и непереваренных остатков пищи.
Хотя животные могут выдерживать кратковременные потери воды, но в целом расход ее должен возмещаться приходом. Потери воды приводят к гибели скорее, чем голодание.
Виды, получающие воду в основном через питье, сильно зависят от наличия водопоев. Это особенно характерно для крупных млекопитающих. В сухих, аридных районах такие животные совершают иногда значительные миграции к водоемам и не могут существовать слишком далеко от них. В африканских саваннах слоны, антилопы, львы, гиены регулярно посещают водопои (рис. 35). Для куланов Бадхызского заповедника водопои определяют летнее размещение табунов, суточный ритм и поведение животных.

Рис. 35. Слоны на водопое
В питьевой воде нуждаются и многие птицы. Ласточки и стрижи пьют на лету, проносясь над поверхностью водоема. Рябки в пустынях ежедневно совершают многокилометровые перелеты к водопоям и приносят воду птенцам. Самцы рябков используют исключительный в своем роде способ переноса воды – они пропитывают ею оперение на груди, а птенцы отжимают клювами набухшие перья.
В то же время многие животные могут обходиться совсем без питьевой воды, получая влагу иными способами.
Влажность воздуха также очень важна для животных, так как от нее зависит величина испарения с поверхности тела (рис. 36). Потери воды через испарение обусловлены также строением покровов. Некоторые виды не могут обитать в сухом воздухе и нуждаются в полном насыщении его водяными парами. Другие без вреда для себя населяют самые засушливые районы.
Среди ряда групп животных можно выделить гигрофилов и ксерофилов, т. е. влаголюбивые и сухолюбивые виды. Промежуточную группу составляют мезофилы. Среди насекомых, например, гигрофильны кровососущие комары, которые активны преимущественно в вечерние и утренние часы, а днем – либо в пасмурную погоду, либо только в тени, под пологом леса, т. е. при повышенной влажности воздуха. Ксерофильны жуки-скакуны, пустынные жуки-чернотелки, пустынная саранча и др.
Способы регуляции водного баланса у животных разнообразнее, чем у растений. Их можно разделить на поведенческие, морфологические и физиологические.
К числу поведенческих приспособлений относятся поиски водопоев, выбор мест обитания, рытье нор и т. п. В норах влажность воздуха приближается к 100 %, даже когда на поверхности очень сухо. Это снижает необходимость испарения через покровы, экономит влагу в организме.
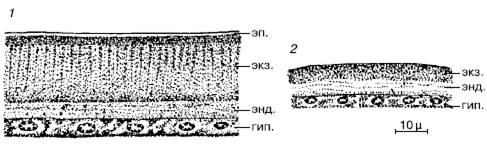
Рис. 36. Строение кутикулы членистоногих (по М. С. Гилярову, 1970):
1– жук мучной хрущах Tenebrio molitor;
2– ветвистоусый рачок Cyclops sp.: гип. – гиподерма, энд. – эндокутикула; экз. – экзокутикула, эп. – эпикутикула
В эффективности поведенческих приспособлений для обеспечения водного баланса можно убедиться на примере пустынных мокриц. Мокрицы – типичные ракообразные, не отличающиеся особыми анатомо-морфологическими приспособлениями к наземному образу жизни. Тем не менее представители рода Hemilepistus освоили самые сухие и жаркие места на Земле – глинистые пустыни. Там они роют глубокие вертикальные норки, где всегда влажно, и покидают их, выходя на поверхность лишь в те часы суток, когда высока влажность приземного слоя воздуха. Когда почва иссушается особенно сильно и возникает угроза снижения влажности воздуха в норке, самки закрывают отверстие сильно склеротизованными передними сегментами тела, создавая замкнутое, насыщенное парами пространство и оберегая молодь от высыхания.
К морфологическим способам поддержания нормального водного баланса относятся образования, способствующие задержанию воды в теле: раковины наземных улиток, ороговевшие покровы рептилий, развитие эпикутикулы у насекомых и т. п. У пустынных жуков-чернотелок срастаются и прирастают к телу надкрылья, вторая пара крыльев редуцируется и между телом и надкрыльями образуется камера, куда выходят дыхальца насекомого. Эта камера открывается наружу лишь небольшой узкой щелью, воздух в ней насыщен водяными парами. Части тела, соприкасающиеся с внешней средой, защищены непроницаемой для воды эпикутикулой.
Физиологические приспособления к регуляции водного обмена – это способность к образованию метаболической влаги, экономии воды при выделении мочи и кала, развитие выносливости к обезвоживанию организма, величина потоотделения и отдачи воды со слизистых.
Выносливость к обезвоживанию, как правило, выше у животных, подвергающихся тепловым перегрузкам. Для человека потеря воды, превышающая 10 % массы тела, смертельна. Верблюды переносят потери воды до 27 %, овцы – до 23, собаки – до 17 %.
Экономия воды в пищеварительном тракте достигается всасыванием воды кишечником и продуцированием сухого кала. Содержание воды в испражнениях животных варьирует в зависимости от состава корма, но в целом отражает приспособленность к обитанию в разных условиях влажности. Например, на 100 г сухого помета коров на пастбище приходится 566 г воды, тогда как у верблюдов – 109, а при безводной диете – всего 76 г.
У насекомых, обитающих в аридных районах, выделительные органы – мальпигиевы сосуды – свободными концами входят в тесный контакт со стенкой задней кишки и всасывают воду из ее содержимого. Таким образом вода вновь возвращается в организм (пустынные жуки-чернотелки, муравьиные львы, личинки божьих коровок и др.).
Для экономии воды, выводимой через почки, нужна перестройка азотного обмена. При распаде белков у большинства водных организмов образуется аммиак, который токсичен для цитоплазмы даже в малых концентрациях. На процесс его образования и выведения тратится много воды. У наземных животных аммиак присутствует среди продуктов обмена только у тех форм, которые обитают в условиях достаточного обеспечения водой, например у тлей, непрерывно питающихся соком растений. Основной компонент выделяемой мочи у наземных млекопитающих – мочевина. Это менее токсичный продукт обмена, который может накапливаться в плазме и полостных жидкостях и выводиться в более концентрированных растворах, что экономит воду. С мочой выводятся также различные соли. Общая концентрация мочи по сравнению с плазмой может служить показателем способности к экономии воды при экскреции. У человека моча концентрированнее плазмы в 4,2 раза, у овец – в 7,6, у верблюда – в 8, у тушканчиков – в 14 раз.
Чешуйчатые пресмыкающиеся и сухопутные черепахи – группы, освоившие наиболее аридные районы, – выделяют малорастворимую мочевую кислоту. Это же характерно для птиц и высших насекомых. Паукообразные выделяют гуанин. При образовании гуанина и мочевой кислоты затрачивается минимальное количество воды.
Жизнь за счет метаболической влаги доступна не всем животным. Окисление жиров требует большого количества кислорода, а дополнительная вентиляция легких в сухом воздухе сопровождается потерей водяных паров. Жир в горбах верблюдов не является для них основным источником водоснабжения, так как расход воды на усиленное дыхание при терморегуляции равен или даже превышает количество получаемой метаболической воды. Поэтому верблюды нуждаются в периодическом питье.
Мелкие млекопитающие, спасающиеся от жары в прохладных норах, могут покрывать значительную часть своих расходов воды в результате окислительных процессов, так как им не требуется дополнительно вода на терморегуляцию. Почти исключительно на сухом корме живут такие пустынные виды, как многие тушканчики, американская кенгуровая крыса, африканская песчанка и др. (рис. 37). Кенгуровых крыс содержали в лаборатории на сухой перловой крупе. При этом из 100 г корма, потребляемого зверьком за месяц, образуется около 54 г воды. Кроме нее, животные использовали лишь абсорбированную крупой влагу, содержание которой, в зависимости от влажности воздуха, составляет от 10 до 18 %.

Рис. 37. Животные, способные жить за счет метаболической влаги: 1 – пустынный мешотчатый прыгун; 2 – карликовая песчанка; 3 – жирнохвостый тушканчик (по В. Е. Соколову и др., 1977)
Метаболическую воду в большей мере, чем позвоночные животные, могут использовать насекомые, так как трахейная система насекомых осуществляет эффективный воздушный дренаж с малыми потерями на испарение. У многих видов жировое тело служит преимущественно источником воды, а не энергетических запасов. Гусеницы платяной моли, мельничной огневки, амбарный и рисовый долгоносики и многие другие живут исключительно за счет сухой пищи.
Испарение, связанное с необходимостью терморегуляции, может служить причиной истощения водных ресурсов организма. В пустынях противостоять перегреву путем испарения воды могут только крупные животные. Общая тепловая нагрузка пропорциональна относительной поверхности и поэтому особенно велика для мелких форм. Для животного массой 100 г расход воды составил бы в час около 15 % от массы тела, а массой 10 г – около 30 %, т. е. за немногие часы была бы истрачена вся вода организма. Поэтому мелкие гомойотермные животные в сухом и жарком климате избегают воздействия жары и экономят влагу, укрываясь под землей.
У пойкилотермных повышение температуры тела вслед за нагреванием воздуха позволяет избегать излишних потерь воды, которая тратится у гомойотермных для поддержания постоянной температуры.
Преимущества колеблющейся температуры тела используют и животные с хорошей температурной регуляцией, специализированные к жизни в пустыне. Например, верблюды способны отключать на некоторое время терморегуляционное испарение. При этом животное массой 500 кг аккумулирует около 10 500 кДж, для рассеивания которых потребовалось бы затратить 5 л воды. Накопленное тепло выводится из организма ночью путем прямого излучения, когда воздух становится прохладнее тела.
Пойкилотермные животные, однако, не могут полностью избежать потерь воды на испарение. Даже у рептилий с их ороговевшим эпидермисом потери воды через кожу значительны. У мелких ящериц они могут достигать 20 % и более от массы тела за сутки. Поэтому и для пойкилотермных основной путь сохранения водного баланса при жизни в пустыне – это избегание излишних тепловых нагрузок.
3.4. Основные пути приспособления живых организмов к условиям среды
Во всем разнообразии приспособлений живых организмов к неблагоприятным условиям среды можно выделить три основных пути.
Активный путь – это усиление сопротивляемости, развитие регуляторных процессов, позволяющих осуществить все жизненные функции организмов, несмотря на отклонения фактора от оптимума. По отношению к температуре, например, этот путь в зачаточной форме проявляется у некоторых высших растений, несколько сильнее развит у пойкилотермных животных, но особенно ярко выражен при гомойотермии. Активное противостояние иссушению особенно характерно для склерофитов среди растений, ксерофильных насекомых (например, пустынных чернотелок), крупных гомойотермных животных аридных районов.
Пассивный путь – подчинение жизненных функций организма изменению факторов среды. При недостатке тепла это приводит к угнетению жизнедеятельности и понижению уровня метаболизма, что способствует экономному использованию энергетических запасов. Компенсаторно повышается устойчивость клеток и тканей организма. Пассивный путь адаптации к влиянию неблагоприятных температур свойствен всем растениям и пойкилотермным животным. Среди млекопитающих и птиц преимущества пассивного приспособления в неблагоприятные периоды года используют гетеротермные виды, впадающие в оцепенение или спячку. Элементы пассивной адаптации присущи и типичным гомойотермным животным, обитающим в условиях крайне низких температур. Это выражается в некотором снижении уровня обмена, замедлении темпов роста и развития, что позволяет экономнее тратить ресурсы по сравнению с быстро развивающимися видами.
Пассивное подчинение водному режиму среды свойственно пойкилогидрическим растениям и животным, способным выносить высыхание: напочвенным водорослям, лишайникам, нематодам, коловраткам и т. п.
Избегание неблагоприятных воздействий – третий возможный путь приспособления к среде. Общий способ для всех групп организмов – выработка таких жизненных циклов, при которых наиболее уязвимые стадии развития завершаются в самые благоприятные по температурным и другим условиям периоды года. Для животных основным способом избегания пессимальных температур являются разнообразные формы поведения. Изменения в ростовых процессах растений – в известной мере экологический аналог поведения животных. Например, карликовость тундровых растений помогает организмам использовать тепло приземного слоя и избегать влияния низких температур воздуха. Растения-эфемероиды в жарких пустынях избегают засухи, успевая отцвести за краткий весенний период.
Избегание, уход от действия крайних температур или недостатка влаги свойствен организмам в той или иной мере и при активном, и при пассивном пути адаптации к среде. Все три пути приспособления характерны и по отношению к другим экологическим факторам среды. Чаще всего приспособление вида к среде осуществляется тем или иным сочетанием всех трех возможных путей адаптаций.
Дата добавления: 2015-11-26; просмотров: 1394;