Глава 3. ВАЖНЕЙШИЕ АБИОТИЧЕСКИЕ ФАКТОРЫ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ 1 страница
Температура
Температура отражает среднюю кинетическую скорость атомов и молекул в какой-либо системе. От температуры зависит и скорость в организме биохимических реакций, составляющих обмен веществ. Повышение температуры увеличивает количество молекул, обладающих энергией активации. Коэффициент, показывающий, во сколько раз изменяется скорость реакций при изменении температуры на 10 °C, обозначают Q10. Для большинства химических реакций величина этого коэффициента равна 2–3 (закон Вант-Гоффа). Изменения температуры приводят также к изменениям стереохимической специфичности макромолекул: третичной и четвертичной структуры белков, строения нуклеиновых кислот, организации мембран и других структур клетки. Так как величина Q10 для разных биохимических реакций различна, то изменения температуры могут сильно нарушить сбалансированность обмена веществ, если скорости сопряженных процессов изменятся различным образом. Сильное понижение температуры вызывает опасность такого замедления обмена веществ, при котором окажется невозможным осуществление основных жизненных функций организма. Критическим моментом является замерзание воды в клетках, так как появление кристалликов льда несовместимо с сохранением целостности внутриклеточных структур. Повышение температуры ведет к денатурации белков, в среднем в области около 60 °C, но рассогласование биохимических и физиологических процессов начинается раньше, уже при некотором превышении 42–43 °C. Излишнее усиление метаболизма при высоких температурах тела также может вывести организм из строя еще задолго до теплового разрушения ферментов, так как резко возрастают потребности в питательных веществах и кислороде, которые далеко не всегда могут быть удовлетворены. Таким образом, жизнь организмов в среде с низкими, высокими и колеблющимися температурами представляет сложную задачу адаптации, решаемую в ходе эволюции и индивидуального развития.
В процессе эволюции у живых организмов выработались разнообразные приспособления, позволяющие регулировать обмен веществ при изменениях температуры окружающей среды. Это достигается двумя путями: 1) различными биохимическими и физиологическими перестройками (изменение набора, концентрации и активности ферментов, обезвоживание, понижение точки замерзания растворов тела и т. д.); 2) поддержанием температуры тела на более стабильном уровне, чем температура окружающей среды, что позволяет не слишком нарушать сложившийся ход биохимических реакций.
3.1.1. Температурные границы существования видов
В среднем активная жизнедеятельность организмов требует довольно узкого диапазона температур, ограниченного критическими порогами замерзания воды и тепловой денатурации белков, примерно в пределах от 0 до +50 °C.Границы оптимальных температур соответственно должны быть еще более узкими. Однако реально эти границы преодолеваются в природе у многих видов за счет специфических адаптаций. Существуют экологические группы организмов, оптимум которых сдвинут в сторону низких или высоких температур.
Криофилы – виды, предпочитающие холод и специализированные к жизни в этих условиях. Свыше 80 % земной биосферы относится к постоянно холодным областям с температурой ниже +5 °C – это глубины Мирового океана, арктические и антарктические пустыни, тундры, высокогорья. Обитающие здесь виды обладают повышенной холодостойкостью. Основные механизмы этих адаптаций биохимические. Ферменты холодолюбивых организмов обладают такими особенностями строения, которые позволяют им эффективно понижать энергию активации молекул и поддерживать клеточный метаболизм при температурах, близких к 0 °C. Большую роль играют также механизмы, предотвращающие образование льда внутри клеток. При этом реализуются два основных пути – противостояние замерзанию (резистентность) и устойчивость к замерзанию (толерантность).
Биохимический путь противостояния замерзанию – накопление в клетках макромолекулярных веществ – антифризов, которые понижают точку замерзания жидкостей тела и препятствуют образованию кристаллов льда в организме. Такого типа холодовые адаптации обнаружены, например, у антарктических рыб семейства нототениевых, которые живут при температуре тела -1,86 °C, плавая под поверхностью сплошного льда в воде с такою же температурой (рис. 11). Мелкая тресковая рыба сайка в Северном Ледовитом океане плавает в водах с температурой не выше +5 °C, а нерестится зимой в переохлажденных водах у побережья. Глубоководные рыбы в приполярных районах все время находятся в переохлажденном состоянии.
Предельная температура, при которой еще возможна активность клеток, зафиксирована у микроорганизмов. В холодильных камерах мясные продукты могут быть испорчены за счет деятельности бактерий при температурах до -10-12 °C. Ниже этих температур роста и развития одноклеточных организмов не происходит.
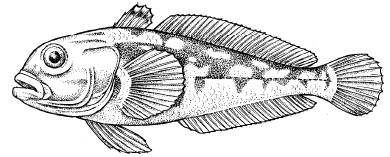
Рис. 11. Антарктическая рыба трематом-пестряк с температурой тела -1,98 °C (см.: Жизнь животных. Т. 4. 1971)
Другой путь холодостойкости – выносливость к замерзанию – связан с временным прекращением активного состояния (гипобиозом или криптобиозом). Образование кристалликов льда внутри клеток необратимо нарушает их ультраструктуру и приводит к гибели. Но многие криофилы способны переносить образование льда во внеклеточных жидкостях. Этот процесс приводит к частичной дегидратации клеток, что повышает их устойчивость. У насекомых накопление защитных органических веществ, таких как глицерин, сорбит, маннит и других, препятствует кристаллизации внутриклеточных растворов и позволяет переживать критические морозные периоды в состоянии оцепенения. Так, жуки-жужелицы в тундрах выдерживают переохлаждение до -35 °C, накапливая к зиме до 25 % глицерина и снижая содержание воды в теле с 65 до 54 %. Летом глицерин в их теле не обнаруживается. Некоторые насекомые выдерживают зимой до -47 и даже -50 °C с замерзанием внеклеточной, но не внутриклеточной влаги. Морские обитатели практически не сталкиваются с температурами ниже -2 °C, но беспозвоночные приливно-отливной зоны (моллюски, усоногие раки и др.) зимой во время отлива переносят замерзание до – (15–20) °С. Клетки под микроскопом выглядят сморщенными, но кристаллов льда в них не обнаруживается. Устойчивость к замерзанию может проявляться и у эвритермных видов, оптимальные температуры развития которых далеки от 0 °C.
Термофилы – это экологическая группа видов, оптимум жизнедеятельности которых приурочен к области высоких температур. Термофилией отличаются многие представители микроорганизмов, растений и животных, встречающихся в горячих источниках, на поверхности прогреваемых почв, в разлагающихся органических остатках при их саморазогревании и т. п.
Верхние температурные пределы активной жизни отличаются у разных групп организмов. Наиболее устойчивы бактерии. У одного из видов архебактерий, распространенных на глубинах вокруг термальных источников («курильщиков»), экспериментально обнаружена способность к росту и делению клеток при температурах, превышающих +110 °C. Некоторые бактерии, окисляющие серу, как, например, Sulfolobus acidocaldarius, размножаются при +(85–90)°С. Обнаружена даже способность ряда видов расти в практически кипящей воде. Естественно, не все бактерии активны при столь высоких температурах, но разнообразие таких видов достаточно велико.
Верхние температурные пороги развития цианобактерий (сине-зеленых водорослей) и других фотосинтезирующих прокариот лежат в более низких пределах от +70 до +73 °C. Термофилы, растущие при +(60–75) °С, есть как среди аэробных, так и анаэробных бактерий, спорообразующих, молочнокислых, актиномицетов, метанообразующих и др. В неактивном состоянии спорообразующие бактерии выдерживают до +200 °C в течение десятков минут, что демонстрирует режим стерилизации предметов в автоклавах.
Термостабильность белков бактерий создается за счет значительного числа малых изменений в их первичной структуре и добавочных слабых связей, определяющих укладку молекул. В транспортных и рибосомных РНК термофилов повышено содержание гуанина и цитозина. Эта пара оснований более термостабильна, чем пара аденин – урацил.
Таким образом, выход температурной устойчивости за пределы средней нормы происходит в основном за счет биохимических адаптаций.
Среди эукариотных организмов – грибов, простейших, растений и животных – также существуют термофилы, но уровень их толерантности к высокой температуре ниже, чем у бактерий. Пределы роста грибного мицелия составляют +(60–62) °С. Известны десятки видов, способных быть активными при +50 °C и выше в таких местообитаниях, как компосты, стога сена, хранящееся зерно, прогреваемая почва, свалки и т. п. Простейшие – амебы и инфузории, одноклеточные водоросли могут размножаться до температуры в +(54–56) °С Высшие растения могут переносить краткосрочные нагревания до +(50–60) °С, но активный фотосинтез даже у пустынных видов тормозится температурами, превышающими +40 °C. Так, в клетках суданской травы при +48 °C движение цитоплазмы останавливается уже через 5 мин. Критические температуры тела некоторых животных, например пустынных ящериц, могут достигать +(48–49) °С, но для большинства видов температуры тела, превышающие +(43–44) °С, несовместимы с жизнью из-за рассогласования физиологических процессов и коагуляции белка коллагена. Таким образом, с усложнением организации живых существ способность их быть активными при высоких температурах понижается.
Узкая специализация и латентные состояния намного раздвигают границы жизни по отношению к отдельным факторам среды. Если средние температурные пределы активности организмов характеризуются диапазоном от 0 до +(40–45) °С, то специализированные виды (криофилы и термофилы) расширяют его более чем вдвое (от -10 до примерно +110 °C), а в состоянии криптобиоза и анабиоза некоторые формы жизни способны выдерживать температуры, близкие к абсолютному нулю или намного превышающие точку кипения виды.
3.1.2. Температура тела и тепловой баланс организмов
Температура тела живых существ по-разному зависит от температуры окружающей среды. Баланс тепла в организме складывается из его прихода и расхода. Источники поступления тепловой энергии делятся на внешние и внутренние. Внешнее, или экзогенное, тепло организм получает от более нагретых воды, воздуха, окружающих предметов, прямой солнечной радиации. При этом большую роль играют площадь покровов и их теплопроводность. Внутреннее, или эндогенное, тепло вырабатывается как обязательный атрибут обмена веществ. Любой организм выделяет в окружающую среду тепло в результате своей жизнедеятельности.
Источником теплообразования в клетках являются два экзотермических процесса: окислительные реакции и расщепление АТФ. Энергия, освобождающаяся при втором процессе, идет, как известно, на осуществление всех рабочих функций клетки, а энергия окисления – на восстановление АТФ. Но и в том, и в другом случае, согласно второму закону термодинамики, часть энергии рассеивается в виде тепла. Тепло, вырабатываемое живыми организмами как побочный продукт биохимических реакций, может служить существенным источником повышения температуры их тела. Общий объем теплопродукции зависит от массы тела и интенсивности метаболизма.
Потери тепла происходят через поверхность тела за счет излучения и теплопроводности, а также за счет энергоемкого испарения воды организмами. По физическим законам на испарение 1 мл воды затрачивается около 539 кал. Соотношение всех этих теплообменных процессов определяет температуру живых существ и влияет на скорость метаболических реакций.
Жизнедеятельность и активность большинства видов на Земле зависят прежде всего от тепла, поступающего извне, а температура тела – от хода внешних температур. Такие организмы называют пойкилотермными. Этот термин обозначает изменчивость теплового режима организмов. Пойкилотермность свойственна всем микроорганизмам, грибам, растениям, беспозвоночным животным и значительной части хордовых. Две группы высших животных – птиц и млекопитающих относят к гомойотермным. Они способны поддерживать постоянную оптимальную температуру тела независимо от температуры среды.
Среди пойкилотермных организмов есть такие, которые всю жизнь проводят в условиях постоянных внешних температур (глубины океанов, пещеры и т. п.), в связи с чем температура их тела не меняется. Такое явление называют ложной гомойотермией (рис. 12). Она свойственна, например, ряду рыб и иглокожих. Среди истинно гомойотермных животных выделяют группу гетеротермных. В нее входят виды, впадающие в спячку или временное оцепенение. Эти виды в активном состоянии поддерживают постоянную температуру тела на высоком уровне, а в неактивном – пониженную, что сопровождается замедлением обмена веществ. Таковы сурки, суслики, летучие мыши, сони, ежи, колибри, стрижи и др. Таким образом, термины «пойкилотермия», «гомойотермия», «ложная гомойотермия» и «гетеротермия» отражают степень изменчивости температуры живых существ.
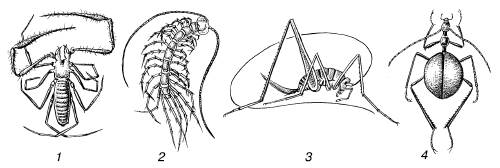
Рис. 12.Ложногомойотермные членистоногие – обитатели пещер: 1– лжескорпион; 2 – многоножка; 3 – кузнечик; 4 – жужелица
Для характеристики организмов по основным источникам используемого тепла используют термины эктотермный и эндотермный. Эктотермия – это жизнь преимущественно за счет нагревания из внешней среды, эндотермия – за счет тепла, вырабатываемого самим организмом.
Масштабы выработки тепла сильно отличаются у разных видов, проявляя зависимость от сложности организации группы, возможностей окислительных реакций, размеров и массы тела, условий среды и других причин. Так, например, бактерии выделяют на грамм веса в час около 450 кал, мухи-дрозофилы – 30, мыши – 8, для человека этот показатель равен 4. В пределах позвоночных животных при сходной массе тела млекопитающие продуцируют в 5–6 раз, а птицы – в 7–8 раз больше тепла, чем рептилии.
Все живые организмы потенциально эндотермны, но сильно различаются по уровню обмена и возможностям сохранения тепла. Нарушения теплового баланса меняют температуру тела. Восстановить нарушенный баланс можно тремя путями: 1) изменением теплопродукции, 2) изменением теплоотдачи и 3) перемещением в пространстве в область предпочитаемых температур. Пойкилотермные и гомойотермные организмы по-разному реализуют возможности температурных адаптаций.
3.1.3. Температурные адаптации пойкилотермных организмов
Температура пойкилотермных изменяется вслед за температурой окружающей среды. Они преимущественно эктотермны, выработки и сохранения собственного тепла у них недостаточно для противостояния тепловому режиму местообитаний. В связи с этим реализуется два основных пути адаптации: специализация и толерантность.
Специализированные виды стенотермны, они приспособлены к жизни в таких участках биосферы, где колебания температур происходят лишь в узких пределах. Выход за эти пределы для них губителен. Например, некоторые одноклеточные водоросли, развивающиеся в горных ледниках на поверхности тающего льда, погибают при температурах, превышающих +(3–5) °С. Растения дождевых тропических лесов не способны переносить снижение температуры до +(5–8) °С. Коралловые полипы живут только в диапазоне температур воды от +20,5 до +30 °C, т. е. в тропическом поясе океана. Голотурия Elpidia glacialis обитает при температуре воды от 0 до +1 °C и не выдерживает отклонения от этого режима ни на один градус.
Другой путь адаптации пойкилотермных видов – развитие устойчивости клеток и тканей к широкому колебанию температур, характерному для большей части биосферы. Этот путь связан с периодическим торможением обмена веществ и перехода организмов в латентное состояние, когда температура среды сильно отклоняется от оптимума.
Эффективные температуры развития пойкилотермных организмов.Зависимость темпов роста и развития от внешних температур дает возможность рассчитать прохождение жизненного цикла видов в конкретных условиях. После холодового угнетения нормальный обмен веществ восстанавливается для каждого вида при определенной температуре, которая называется температурным порогом развития, или биологическим нулем развития. Чем больше температура среды превышает пороговую, тем интенсивнее протекает развитие и, следовательно, тем скорее завершается прохождение отдельных стадий и всего жизненного цикла организма (рис. 13).
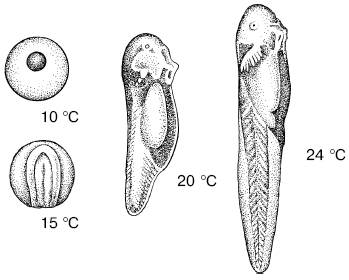
Рис. 13. Состояние развивающихся при разных температурах головастиков через 3 дня после оплодотворения яйца (по С. А. Зернову, 1949)
Для осуществления генетической программы развития пой-килотермным организмам необходимо получить извне определенное количество тепла. Это тепло измеряется суммой эффективных температур. Под эффективной температурой понимают разницу между температурой среды и температурным порогом развития организмов. Для каждого вида она имеет верхние пределы, так как слишком высокие температуры уже не стимулируют, а тормозят развитие.
И порог развития, и сумма эффективных температур для каждого вида свои. Они зависят от исторической приспособленности к условиям жизни. Для семян растений умеренного климата, например гороха, клевера, порог развития низкий: их прорастание начинается при температуре почвы от 0 до +1 °C; более южные культуры – кукуруза и просо – начинают прорастать только при +(8-10) °С, а семенам финиковой пальмы для начала развития нужно прогревание почвы до +30 °C.
Сумму эффективных температур рассчитывают по формуле
X = (T – C) · t,
где X– сумма эффективных температур; T – температура окружающей среды, С – температура порога развития и t – число часов или дней с температурой, превышающей порог развития.
Зная средний ход температур в каком-либо районе, можно рассчитать появление определенной фазы или число возможных генераций интересующего нас вида. Так, в климатических условиях Северной Украины может выплодиться лишь одна генерация бабочки яблонной плодожорки, а на юге Украины – до трех, что необходимо учитывать при разработке мер защиты садов от вредителей. Сроки цветения растений зависят от того, за какой период они набирают сумму необходимых температур. Для зацветания мать-и-мачехи под Петербургом, например, сумма эффективных температур равна 77, кислицы – 453, земляники – 500, а желтой акации – 700 °C.
Сумма эффективных температур, которую нужно набрать для завершения жизненного цикла, часто ограничивает географическое распространение видов. Например, северная граница лесной растительности приблизительно совпадает с июльскими изотермами +(10–12) °С. Севернее тепла для развития деревьев уже не хватает, и зона лесов сменяется безлесными тундрами.
Расчеты эффективных температур необходимы в практике сельского и лесного хозяйства, при борьбе с вредителями, интродукции новых видов и т. п. Они дают первую, приближенную основу для составления прогнозов. Однако на распространение и развитие организмов влияет множество других факторов, поэтому в действительности температурные зависимости оказываются более сложными.
Температурная компенсация.Ряд пойкилотермных видов, обитающих в условиях переменных температур, развивает возможность поддерживать более или менее постоянный уровень обмена веществ в довольно широких пределах изменения температуры тела. Это явление называется температурной компенсацией и происходит в основном за счет биохимических адаптаций. Например, у моллюсков на побережье Баренцева моря, таких, как брюхоногие литторины (Littorina littorea) и двустворчатые мидии (Mytilus edulis), интенсивность обмена, оцениваемая по потреблению кислорода, почти не зависит от температуры в тех пределах, с которыми моллюски встречаются ежедневно во время приливов и отливов. В весенне-летний период этот диапазон достигает более 20 °C (от +6 до +30 °C), и в холодной воде их метаболизм столь же интенсивен, как в теплом воздухе. Это обеспечивается действием ферментов, которые при понижении температуры меняют свою конфигурацию таким образом, что возрастает их сродство к субстрату и реакции протекают более активно.
Другие способы температурной компенсации связаны с заменой действующих ферментов сходными по функции, но работающими при иной температуре (изоферментами). Такие адаптации требуют времени, поскольку происходит инактивация одних генов и включение других с последующими процессами сборки белков. Подобная акклимация (сдвиг температурного оптимума) лежит в основе сезонных перестроек, а также обнаруживается у представителей широко распространенных видов в разных по климату частях ареала. Например, у одного из видов бычков из Атлантического океана в низких широтах Q10 имеет невысокое значение, а в холодных северных водах возрастает при низких температурах и снижается при средних. Результатом этих компенсаций является то, что животные могут поддерживать относительное постоянство активности, так как даже незначительное повышение температуры у критических точек усиливает обменные процессы. Температурные компенсации для каждого вида возможны лишь в определенном диапазоне температур, но не выше и не ниже этой области.
Биохимические адаптации при всей их эффективности не представляют главный механизм противостояния неблагоприятным условиям. На самом деле они являются часто «крайним средством» и эволюционно вырабатываются у видов лишь тогда, когда невозможны другие способы, физиологические, морфо-анатомические или поведенческие, избегать экстремальных воздействий без перестройки основного химизма клеток. Ряд пойкилотермных организмов обладает возможностями частичной регуляции теплообмена, т. е. некоторыми способами увеличить поступление тепла в организм или отвести его избыток. В основном эти адаптации возникают у многоклеточных растений или животных и в каждой группе имеют свою специфику.
Элементы регуляции температуры у растений.Растения вырабатывают мало метаболического тепла вследствие эффективного перевода химической энергии из одних форм в другие, поэтому эндотермия не может быть использована ими для терморегуляции. Будучи организмами прикрепленными, они должны существовать при том тепловом режиме, который создается в местах их произрастания. Однако совпадение температур тела растения и среды скорее надо считать исключением, чем правилом, из-за разницы скоростей поступления и отдачи тепла. Высшие растения умеренно холодного и умеренно теплого поясов эвритермны. Тепловой режим растений весьма изменчив. Температура разных органов различна в зависимости от их расположения относительно падающих лучей и разных по степени нагретости слоев воздуха (рис. 14). Тепло поверхности почвы и приземного слоя воздуха особенно важно для тундровых и высокогорных растений. Приземистость, шпалерные и подушковидные формы роста, прижатость листьев розеточных и полурозеточных побегов к субстрату у арктических и высокогорных растений можно рассматривать как адаптацию к лучшему использованию тепла в условиях, где его мало (рис. 15).
А, Б
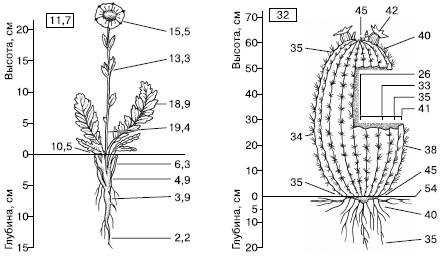
Рис. 14. Температура (в °С) разных органов растений (из В. Лархера, 1978).
В рамках дана температура воздуха на высоте растения:
A – растение тундры Novosieversia glacialis,
Б – кактус Ferocactus wislisenii

Рис. 15. Высокогорное растение Копетдага качим подушковидный – Gypsophila aretiodes (по К. П. Попову, Э. М. Сейфулину, 1994)
В дни с переменной облачностью надземные органы растений испытывают резкие перепады температуры. Например, у дубравного эфемероида пролески сибирской, когда облака закрывают солнце, температура листьев может упасть с +(25–27) °С до +(10–15) °С, а затем, когда растения снова освещаются солнцем, поднимается до прежнего уровня. В пасмурную погоду температура листьев и цветков близка к температуре окружающего воздуха, но чаще бывает на несколько градусов ниже из-за транспирации. У многих растений разница температур заметна даже в пределах одного листа. Обычно верхушка и края листьев холоднее, поэтому при ночном охлаждении в этих местах в первую очередь конденсируется роса и образуется иней. При нагревании солнечными лучами температура растения может быть значительно выше температуры окружающего воздуха. Иногда эта разница доходит более чем до 20 °C, как, например, у крупных мясистых стеблей пустынных кактусов или стволов одиночно стоящих деревьев.
Основное средство отведения избытка тепла и предотвращения ожогов – устьичная транспирация. Испарение 1 г воды выводит из тела растения около 583 кал (2438 Дж). Если в жаркую солнечную погоду смазать вазелином ту поверхность листа, на которой расположены устьица, лист очень быстро гибнет от перегрева и ожогов. Усиление транспирации при повышении температуры среды охлаждает растение. Однако этот механизм терморегуляции эффективен лишь в условиях достаточного водообеспечения, что редко бывает в аридных районах.
Растения обладают также рядом морфологических адаптации, направленных на предотвращение перегрева. Этому служат густая опушенность листьев, рассеивающая часть солнечных лучей, глянцевитая поверхность, способствующая их отражению, уменьшение поглощающей лучи поверхности. Многие злаки, как, например, ковыль или овсяница, в жару свертывают листовые пластинки в трубочку, у эвкалиптов листья располагаются ребром к солнечным лучам, у части растений аридных районов листва полностью или частично редуцируется (саксаулы, кактусы, кактусовидные молочаи и др.).
В экстремально холодных условиях средствами получения дополнительного тепла служат также некоторые морфологические особенности растений. Основные из них – особые формы роста. Карликовость и образование стелющихся форм позволяет использовать микроклимат приземного слоя летом и быть защищенными снеговым покровом зимой. Своеобразны растения-подушки. Их полусферическая форма создается за счет густого ветвления и слабого роста побегов. Листья располагаются лишь на периферии, в результате чего экономится общая поверхность растения, через которую происходит рассеивание тепла. Как известно, из всех геометрических фигур у шара наименьшее отношение поверхности к объему, что и реализуется в форме растения. Значительная часть холодостойких растений имеет темную окраску, что помогает лучше поглощать тепловые лучи и нагреваться даже под снегом. В Антарктиде летом температура темно-коричневых лишайников бывает выше 0 °C даже под слоем снега в 30 см.
И транспирация, и морфологические адаптации, направленные на поддержание теплового баланса растений, подчиняются физическим законам природы и относятся к способам физической терморегуляции. У растений физическая терморегуляция хотя и представлена различными элементами, но в целом эффективность ее низка и распространяется лишь на несколько процентов общего теплового потока через организмы. Эти элементы терморегуляции позволяют растениям выживать в условиях, когда температура среды приближается к основным критическим значениям, но не могут стабилизировать их общий тепловой баланс. Более существенное значение для растений имеют физиологические механизмы температурных адаптации, повышающие их толерантность к холоду или перегреву (накопление в клетках антифризов, листопад, отмирание надземных частей, уменьшение в клетках воды и т. п.).
В разные фазы онтогенеза требования к теплу различны. В умеренном поясе прорастание семян происходит обычно при более низких температурах, чем цветение, а для цветения требуется более высокая температура, чем для созревания плодов.
По степени адаптации растений к условиям крайнего дефицита тепла можно выделить три группы:
1) нехолодостойкие растения– сильно повреждаются или гибнут при температурах, еще не достигающих точки замерзания воды. Гибель связана с инактивацией ферментов, нарушением обмена нуклеиновых кислот и белков, проницаемости мембран и прекращением тока ассимилятов. Это растения дождевых тропических лесов, водоросли теплых морей;
2) неморозостойкие растения– переносят низкие температуры, но гибнут, как только в тканях начинает образовываться лед. При наступлении холодного времени года у них повышается концентрация осмотически активных веществ в клеточном соке и цитоплазме, что понижает точку замерзания до – (5–7) °С. Вода в клетках может охлаждаться ниже точки замерзания без немедленного образования льда. Переохлажденное состояние неустойчиво и длится чаще всего несколько часов, что, однако, позволяет растениям переносить заморозки. Таковы некоторые вечнозеленые субтропические растения – лавры, лимоны и др.;
3) льдоустойчивые, или морозоустойчивые, растения– произрастают в областях с сезонным климатом, с холодными зимами. Во время сильных морозов надземные органы деревьев и кустарников промерзают, но тем не менее сохраняют жизнеспособность, так как в клетках кристаллического льда не образуется. Растения подготавливаются к перенесению морозов постепенно, проходя предварительную закалку после того, как заканчиваются ростовые процессы. Закалка заключается в накоплении в клетках сахаров (до 20–30 %), производных углеводов, некоторых аминокислот и других защитных веществ, связывающих воду. При этом морозоустойчивость клеток повышается, так как связанная вода труднее оттягивается образующимися во внеклеточных пространствах кристаллами льда.
Дата добавления: 2015-11-26; просмотров: 1270;