Глава 3. ВАЖНЕЙШИЕ АБИОТИЧЕСКИЕ ФАКТОРЫ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ 3 страница
Видимая радиация несет приблизительно 50 % суммарной энергии. С областью видимой радиации, воспринимаемой человеческим глазом, почти совпадает ФР – физиологическая радиация (длина волны 300–800 нм), в пределах которой выделяют ФАР – область фотосинтетически активной радиации (380–710 нм). Область ФР можно условно разделить на ряд зон: ультрафиолетовую (менее 400 нм), сине-фиолетовую (400–500 нм), желто-зеленую (500–600 нм), оранжево-красную (600–700 нм) и дальнюю красную (более 700 нм).
Видимый свет для фототрофных и гетеротрофных организмов имеет разное экологическое значение.
Зеленым растениям свет нужен для образования хлорофилла, формирования гранальной структуры хлоропластов; он регулирует работу устьичного аппарата, влияет на газообмен и транспирацию, активизирует ряд ферментов, стимулирует биосинтез белков и нуклеиновых кислот. Свет влияет на деление и растяжение клеток, ростовые процессы и на развитие растений, определяет сроки цветения и плодоношения, оказывает формообразующее воздействие. Но самое большое значение имеет свет в осуществлении процесса фотосинтеза. С этим связаны основные адаптации растений по отношению к свету.
Фотоавтотрофы способны ассимилировать СО2, используя лучистую энергию Солнца и преобразуя ее в энергию химических связей в органических соединениях. Пурпурные и зеленые бактерии, имеющие бактериохлорофиллы, способны поглощать свет в длинноволновой части (максимумы в области 800-1100 нм). Это позволяет им существовать даже при наличии только невидимых инфракрасных лучей. Водоросли и высшие зеленые растения поглощают свет в диапазоне, близком к видимому человеческим глазом.
Водоросли обитают в водоемах, но встречаются и на суше на поверхности разных предметов – на стволах деревьев, на заборах, на скалах, на снегу, на поверхности почвы и в ее толще.
В почве находят водоросли на глубине до 2,7 м, но большая часть их обитает в самых верхних слоях (до 1 см). Здесь они являются типичными фототрофами, однако в глубине почвы, в полной темноте, могут переходить на гетеротрофное питание.
В Мировом океане водоросли обитают в освещаемой зоне. Глубже всех проникают красные водоросли. Чаще они обитают на глубинах до 20–40 м, но если прозрачность воды велика, то встречаются до 100 и даже 200 м.
На суше для высших фотоавтотрофных растений условия освещения практически везде благоприятны, и они растут повсюду, где позволяют климатические и почвенные условия, приспосабливаясь к световому режиму данного местообитания.
3.2.2. Экологические группы растений по отношению к свету и их адаптивные особенности
Световой режим любого местообитания определяется интенсивностью прямого и рассеянного света, количеством света (годовой суммарной радиацией), его спектральным составом, а также альбедо – отражательной способностью поверхности, на которую падает свет.
Перечисленные элементы светового режима очень переменчивы и зависят от географического положения, высоты над уровнем моря, от рельефа, состояния атмосферы, характера земной поверхности, растительности, от времени суток, сезона года, солнечной активности и глобальных изменений в атмосфере.
У растений возникают различные морфологические и физиологические адаптации к световому режиму местообитаний.
По требованию к условиям освещения принято делить растения на следующие экологические группы:
1) светолюбивые (световые), или гелиофиты, – растения открытых, постоянно хорошо освещаемых местообитаний;
2) тенелюбивые (теневые), или сциофиты, – растения нижних ярусов тенистых лесов, пещер и глубоководные растения; они плохо переносят сильное освещение прямыми солнечными лучами;
3) теневыносливые, или факультативные гелиофиты, – могут переносить большее или меньшее затенение, но хорошо растут и на свету; они легче других растений перестраиваются под влиянием изменяющихся условий освещения.
Можно отметить некоторые общие приспособительные особенности, свойственные растениям каждой экологической группы.
Световые адаптации гелиофитов и сциофитов.Гелиофиты часто имеют побеги с укороченными междоузлиями, сильно ветвящиеся, нередко розеточные. Листья гелиофитов обычно мелкие или с рассеченной листовой пластинкой, с толстой наружной стенкой клеток эпидермы, нередко с восковым налетом или густым опушением, с большим числом устьиц на единицу площади, часто погруженных, с густой сетью жилок, с хорошо развитыми механическими тканями. У ряда растений листья фотометричные, т. е. повернуты ребром к полуденным лучам или могут менять положение своих частей в зависимости от высоты стояния Солнца. Так, у степного растения софоры листочки непарноперистосложного листа в жаркий день подняты вверх и сложены, у василька русского так же ведут себя сегменты перисторассеченного листа.
Оптический аппарат гелиофитов развит лучше, чем у сциофитов, имеет большую фотоактивную поверхность и приспособлен к более полному поглощению света. Обычно у них лист толще, клетки эпидермы и мезофилла мельче, палисадная паренхима двухслойная или многослойная (у некоторых саван-ных растений Западной Африки – до 10 слоев), нередко развита под верхней и нижней эпидермой. Мелкие хлоропласты с хорошо развитой гранальной структурой в большом числе (до 200 и более) расположены вдоль продольных стенок.
Хлорофилла на сухую массу в листьях гелиофитов приходится меньше, но зато в них содержится больше пигментов I пигментной системы и хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 5: 1. Отсюда высокая фотосинтетическая способность гелиофитов. Компенсационная точка лежит в области более высокой освещенности. Интенсивность фотосинтеза достигает максимума при полном солнечном освещении. У особой группы растений – гелиофитов, у которых фиксация СО2 идет путем С4-дикарбоновых кислот, световое насыщение фотосинтеза не достигается даже при самой сильной освещенности. Это растения из засушливых областей (пустынь, саванн). Особенно много С4-растений среди семейств мятликовых, осоковых, аизовых, портулаковых, амарантовых, маревых, гвоздичных, молочайных. Они способны к вторичной фиксации и реутилизации СО2, освобождающегося при световом дыхании, и могут фотосинтезировать при высоких температурах и при закрытых устьицах, что часто наблюдается в жаркие часы дня.
Обычно С4-растения, особенно сахарный тростник и кукуруза, отличаются высокой продуктивностью.
Сциофиты– это растения, постоянно находящиеся в условиях сильного затенения. При освещенности 0,1–0,2 % могут расти только мхи и селягинеллы. Плауны довольствуются 0,25-0,5 % полного дневного света, а цветковые растения встречаются обычно там, где освещенность в пасмурные дни достигает не менее 0,5–1% (бегонии, недотрога, травы из семейств имбирные, мареновые, коммелиновые).
В северных широколиственных и темнохвойных лесах полог сомкнутого древостоя может пропускать всего 1–2% ФАР, изменяя ее спектральный состав. Сильнее всего поглощаются синие и красные лучи, и пропускается относительно больше желто-зеленых лучей, дальних красных и инфракрасных. Слабая освещенность сочетается с повышенной влажностью воздуха и повышенным содержанием в нем СО2, особенно у поверхности почвы. Сциофиты этих лесов – зеленые мхи, плауны, кислица обыкновенная, грушанки, майник двулистный и др.
Листья у сциофитов располагаются горизонтально, нередко хорошо выражена листовая мозаика. Листья темно-зеленые, более крупные и тонкие. Клетки эпидермы крупнее, но с более тонкими наружными стенками и тонкой кутикулой, часто содержат хлоропласты. Клетки мезофилла крупнее, палисадная паренхима однослойная или имеет нетипичное строение и состоит не из цилиндрических, а из трапециевидных клеток. Площадь жилок вдвое меньше, чем у листьев гелиофитов, число устьиц на единицу площади меньше. Хлоропласты крупные, но число их в клетках невелико.
У сциофитов по сравнению с гелиофитами меньше хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 3: 2. С меньшей интенсивностью протекают у них такие физиологические процессы, как транспирация, дыхание. Интенсивность фотосинтеза, быстро достигнув максимума, перестает возрастать при усилении освещенности, а на очень ярком свету может даже понизиться.
У лиственных теневыносливых древесных пород и кустарников (дуба черешчатого, липы сердцевидной, сирени обыкновенной и др.) листья, расположенные по периферии кроны, имеют структуру, сходную со структурой листьев гелиофитов, и называются световыми, а в глубине кроны – теневые листья с теневой структурой, сходной со структурой листьев сциофитов (рис. 24).
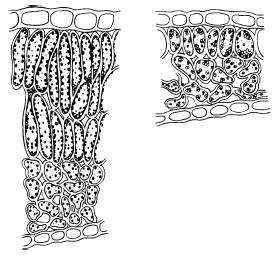
Рис. 24. Поперечный срез светового (слева) и теневого (справа) листа сирени (по И. С. Михайловской, 1977)
Факультативные гелиофиты, или теневыносливые растения, в зависимости от степени теневыносливости имеют приспособительные особенности, сближающие их то с гелиофитами, то со сциофитами. К этой группе можно отнести некоторые луговые растения, лесные травы и кустарники, растущие и в затененных участках леса, и на лесных полянах, опушках, вырубках. На осветленных местах они разрастаются часто сильнее, однако оптимальное использование ФАР у них происходит не при полном солнечном освещении.
У деревьев и кустарников теневая или световая структура листа часто определяется условиями освещения предыдущего года, когда закладываются почки: если закладка почек идет на свету, то формируется световая структура, и наоборот.
Если в одном и том же местообитании закономерно периодически изменяется световой режим, растения в разные сезоны могут проявлять себя то как светолюбивые, то как теневыносливые.
Весной в дубравах под полог леса проникает 50–60 % солнечной радиации. Листья розеточных побегов сныти обыкновенной имеют световую структуру и отличаются высокой интенсивностью фотосинтеза. В это время они создают основную часть органического вещества годичной продукции. Листья сныти летней генерации, появляющиеся при развитом древесном пологе, под который проникает в среднем 3,5 % солнечной радиации, имеют типичную теневую структуру, и интенсивность фотосинтеза их значительно ниже, в 10–20 раз. Подобную двойственность по отношению к свету проявляет и осока волосистая, светолюбивая весной и теневыносливая летом. По-видимому, это свойственно и другим растениям дубравного широкотравья.
Отношение к световому режиму меняется у растений и в онтогенезе. Проростки и ювенильные растения многих луговых видов и древесных пород более теневыносливы, чем взрослые особи.
Иногда у растений меняются требования к световому режиму, когда они оказываются в иных климатических и эдафических условиях. Так, обычные теневыносливые растения хвойного леса – черника, седмичник европейский и некоторые другие – в тундре приобретают особенности гелиофитов.
Наиболее общая адаптация растений к максимальному использованию ФАР – пространственная ориентация листьев. При вертикальном расположении листьев, как, например, у многих злаков и осок, солнечный свет полнее поглощается в утренние и вечерние часы – при более низком стоянии солнца. При горизонтальной ориентации листьев полнее используются лучи полуденного солнца. При диффузном расположении листьев в разных плоскостях солнечная радиация в течение дня утилизируется наиболее полно. Обычно при этом листья нижнего яруса на побеге отклонены горизонтально, среднего направлены косо вверх, а верхнего располагаются почти вертикально.
Считают, что кукуруза является одной из самых высокопродуктивных сельскохозяйственных культур потому, что наряду с высоким КПД фотосинтеза у нее наблюдается диффузное расположение листьев, при котором полнее поглощается ФАР.
На севере, где высота стояния солнца меньше, встречается больше растений с вертикальным расположением листьев, а на юге – с горизонтальным. Для получения большей биомассы выгодны также посевы и насаждения, в которых сочетаются растения с разной пространственной ориентацией листьев, причем в верхнем ярусе лучше иметь растения с вертикальным расположением листьев, которые полнее используют свет при низком стоянии солнца, не препятствуют прохождению полуденных лучей к расположенным в нижнем ярусе листьям с горизонтальной ориентацией.
3.2.3. Свет как условие ориентации животных
Для животных солнечный свет не является таким необходимым фактором, как для зеленых растений, поскольку все гете-ротрофы в конечном счете существуют за счет энергии, накопленной растениями. Тем не менее и в жизни животных световая часть спектра солнечного излучения играет важную роль. Разные виды животных нуждаются в свете определенного спектрального состава, интенсивности и длительности освещения. Отклонения от нормы подавляют их жизнедеятельность и приводят к гибели. Различают виды светолюбивые (фотофилы) и тенелюбивые (фотофобы); эврифотные, выносящие широкий диапазон освещенности, и стенофотные, переносящие узкоограниченные условия освещенности.
Свет для животных необходимое условие видения, зрительной ориентации в пространстве. Рассеянные, отраженные от окружающих предметов лучи, воспринимаемые органами зрения животных, дают им значительную часть информации о внешнем мире. Развитие зрения у животных шло параллельно с развитием нервной системы.
Полнота зрительного восприятия окружающей среды зависит у животных в первую очередь от степени эволюционного развития. Примитивные глазки многих беспозвоночных – это просто светочувствительные клетки, окруженные пигментом, а у одноклеточных – светочувствительный участок цитоплазмы. Процесс восприятия света начинается с фотохимических изменений молекул зрительных пигментов, после чего возникает электрический импульс. Органы зрения из отдельных глазков не дают изображения предметов, а воспринимают только колебания освещенности, чередование света и тени, свидетельствующие об изменениях в окружающей среде. Образное видение возможно только при достаточно сложном устройстве глаза. Пауки, например, могут различать контуры движущихся предметов на расстоянии 1–2 см. Наиболее совершенные органы зрения – глаза позвоночных, головоногих моллюсков и насекомых. Они позволяют воспринимать форму и размеры предметов, их цвет, определять расстояние.
Способность к объемному видению зависит от угла расположения глаз и от степени перекрывания их полей зрения. Объемное зрение, например, характерно для человека, приматов, ряда птиц – сов, соколов, орлов, грифов. Животные, у которых глаза расположены по бокам головы, имеют монокулярное, плоскостное зрение.
Предельная чувствительность высокоразвитого глаза огромна. Привыкший к темноте человек может различить свет, интенсивность которого определяется энергией всего пяти квантов, что близко к физически возможному пределу.
Понятие видимого света в некоторой мере условно, так как отдельные виды животных сильно различаются по способности воспринимать разные лучи солнечного спектра. Для человека область видимых лучей – от фиолетовых до темно-красных.
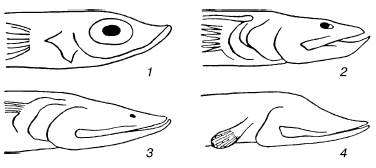
Рис. 25. Разная степень редукции глаз у глубоководных рыб семейства Scopelidae (по Ф. Швердпфегеру, 1963):
1– Chlorophthalmus productus (с глубины 750 м);
2– Bathypterois dubius (800-1000 м);
3– Benthosaurus grallator (3000 м);
4– Dathymicrops regis (5000 м)
Некоторые животные, например гремучие змеи, видят инфракрасную часть спектра и ловят добычу в темноте, ориентируясь при помощи органов зрения. Для пчел видимая часть спектра сдвинута в более коротковолновую область. Они воспринимают как цветовые значительную часть ультрафиолетовых лучей, но не различают красных.
Кроме эволюционного уровня группы, развитие зрения и его особенности зависят от экологической обстановки и образа жизни конкретных видов (рис. 25, 26).
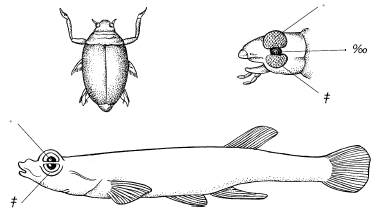
Рис. 26. Глаза жука-вертячки Gyrinus и четырехглазой рыбки Anableps, плавающих у поверхности воды:
а– верхняя часть глаза, приспособленная к зрению в воздухе;
б– нижняя часть глаза, приспособленная к зрению в воде;
в– место прикрепления усика у вертячки
У постоянных обитателей пещер, куда не проникает солнечный свет, глаза могут быть полностью или частично редуцированы, как, например, у слепых жуков жужелиц, протеев среди амфибий и др.
Способность к различению цвета в значительной мере зависит и от того, при каком спектральном составе излучения существует или активен вид. Большинство млекопитающих, ведущих происхождение от предков с сумеречной и ночной активностью, плохо различают цвета и видят все в черно-белом изображении (собачьи, кошачьи, хомяки и др.). Такое же зрение характерно для ночных птиц (совы, козодои). Дневные птицы имеют хорошо развитое цветовое зрение.
Жизнь при сумеречном освещении приводит часто к гипертрофии глаз. Огромные глаза, способные улавливать ничтожные доли света, свойственны ведущим ночной образ жизни лемурам, обезьянам лори, долгопятам, совам и др.
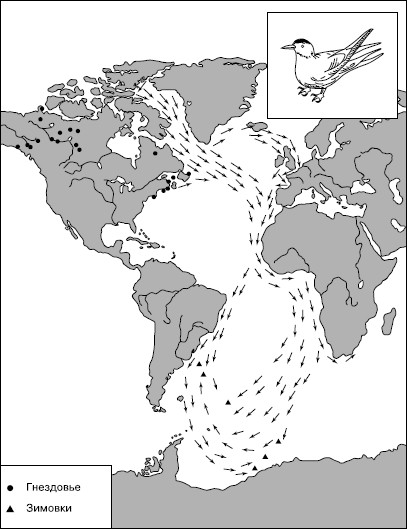
Рис. 27. Пролет североамериканских полярных крачек на зимовки (по И. Штейнбахеру, 1956)
Животные ориентируются с помощью зрения во время дальних перелетов и миграций. Птицы с поразительной точностью выбирают направление полета, преодолевая иногда тысячи километров от гнездовий до мест зимовок (рис. 27).
Доказано, что при таких дальних перелетах птицы хотя бы частично ориентируются по солнцу и звездам, т. е. астрономическим источникам света. При вынужденном отклонении от курса они способны к навигации, т. е. к изменению ориентации, чтобы попасть в нужную точку Земли. При неполной облачности ориентация сохраняется, если видна хотя бы часть неба. В сплошной туман птицы не летят или, если он застает их в пути, продолжают лететь вслепую и часто сбиваются с курса.
Способность птиц к навигации доказана многими опытами.
Птицы, сидящие в клетках, в состоянии предмиграционного беспокойства всегда ориентируются в сторону зимовок, если они могут наблюдать за положением солнца или звезд. Например, когда чечевиц перевезли с побережья Балтийского моря в Хабаровск, они изменили свою ориентацию в клетках с юго-восточной на юго-западную. Зимуют эти птицы в Индии. Таким образом, они способны правильно выбирать направление полета на зимовку из любой точки Земли. Днем птицы учитывают не только положение солнца, но и смещение его в связи с широтой местности и временем суток. Опыты в планетарии показали, что ориентация птиц в клетках меняется, если менять перед ними картину звездного неба в соответствии с направлением предполагаемого перелета.
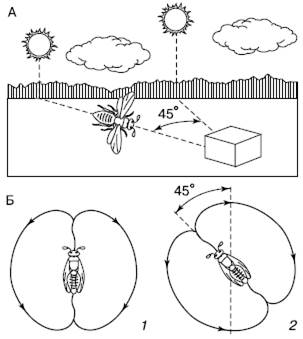
Рис. 28. Ориентация полета пчел по положению солнца (по W. Jacobs, М. Renner, 1974):
A – ориентация полета за взятком;
Б – танец пчелы-разведчицы на вертикальных сотах:
1– положение оси «восьмерки» в случае, если направление к месту взятка совпадает с направлением на солнце;
2– отклонение оси «восьмерки» при передвижении солнца
Навигационная способность птиц врожденная. Она не приобретается за счет жизненного опыта, а создается естественным отбором как система инстинктов. Точные механизмы такой ориентации еще плохо изучены. Гипотеза ориентации птиц в перелетах по астрономическим источникам света в настоящее время подкреплена материалами опытов и наблюдений.
Способность к подобного рода ориентации свойственна и другим группам животных. Среди насекомых она особенно развита у пчел. Пчелы, нашедшие нектар, передают другим информацию о том, куда лететь за взятком, используя в качестве ориентира положение солнца. Пчела-разведчица, открывшая источник корма, возвращается в улей и начинает на сотах танец, совершая быстрые повороты. При этом она описывает фигуру в виде восьмерки, поперечная ось которой наклонена по отношению к вертикали. Угол наклона соответствует углу между направлениями на солнце и на источник корма (рис. 28). Когда медосбор очень обилен, разведчицы сильно возбуждены и могут танцевать долго, в течение многих часов, указывая сборщицам путь к нектару. За время их танца угол наклона восьмерки постепенно смещается в соответствии с движением солнца по небу, хотя пчелы в темном улье и не видят его. Если солнце скрывается за облаками, пчелы ориентируются на поляризованный свет свободного участка неба. Плоскость поляризации света зависит от положения солнца.
Влажность
Протекание всех биохимических процессов в клетках и нормальное функционирование организма в целом возможны только при достаточном обеспечении его водой – необходимым условием жизни. Поддержание водного баланса имеет огромное значение для всех живых организмов.
Проблемы водообеспечения особенно важны для обитателей суши. Особенности поддержания водного баланса зависят от того, в какой экологической обстановке они живут, какой образ жизни ведут, насколько могут использовать различные источники влаги и задерживать воду в теле.
3.3.1. Адаптация растений к поддержанию водного баланса
Низшие наземные растения из влажного субстрата поглощают воду погруженными в него частями таллома, а влагу дождя, росы и тумана – всей поверхностью. В максимально набухшем состоянии лишайники содержат в 20–30 раз больше воды, чем сухого вещества.
Среди высших наземных растений мохообразные поглощают воду из почвы ризоидами, а большинство других – корнями, специализированными органами, всасывающими воду. В клетках корня развивается сосущая сила чаще всего в несколько атмосфер, но этого достаточно для извлечения из почвы большей части связанной воды. Лесные деревья умеренной зоны развивают сосущую силу корней около 3 · 106 Па (30 атм), некоторые травянистые растения (земляника лесная, медуница неясная) – до 2 · 106 (20 атм) и даже свыше 4 · 106 Па (40 атм) (смолка обыкновенная); растения сухих областей – до 60 атм.
Когда в непосредственной близости от корней запасы воды в почве истощаются, корни растут в направлении большей влажности, так что корневая система растений постоянно находится в движении. У степных и пустынных растений часто можно видеть эфемерные корни, быстро вырастающие в периоды увлажнения почвы, а с наступлением засушливого периода засыхающие.
По типу ветвления различают следующие корневые системы:
1) экстенсивная охватывает большой объем почвы, но сравнительно слабо ветвится, так что почва пронизана корнями негусто. Таковы корневые системы у многих степных и пустынных растений (саксаула, верблюжьей колючки), у деревьев умеренной полосы (сосны обыкновенной, березы повислой), а из трав у люцерны серповидной, василька шероховатого и др.;
2) интенсивная– охватывает сравнительно небольшой объем почвы, но густо пронизывает ее многочисленными сильно ветвящимися корнями, как, например, у степных дерновинных злаков (ковылей, типчака и др.), у ржи, пшеницы. Между этими типами корневых систем есть переходные.
Корневые системы очень пластичны и резко реагируют на изменение условий, в первую очередь увлажнения (рис. 29). При недостатке влаги корневая система становится экстенсивнее. Так, при выращивании ржи в разных условиях общая длина корней (без корневых волосков) в 1000 см3 почвы варьирует от 90 м до 13 км, а поверхность корневых волосков может увеличиться в 400 раз.
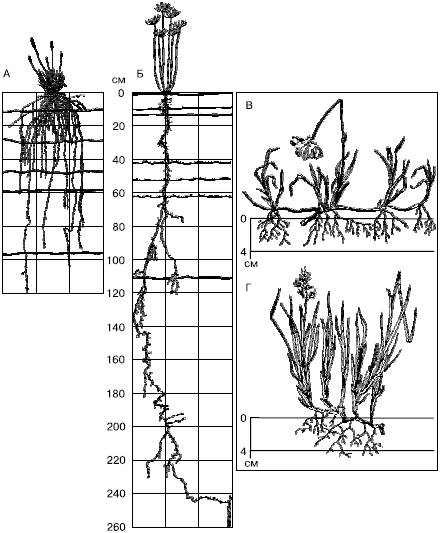
Рис. 29. Корневые системы стенных и тундровых растений (по М. С. Шалыту и Б. А. Тихомирову, 1963):
А – Festuca sulcata;
Б – Euphorbia gerardiana на черноземах в Аскания-Нова;
В – Eriophorum scheuchzeri;
Г – Hierochloe alpina – из тундр Таймыра
Всасывание воды корнями затруднено при большой сухости почвы, засолении или сильной кислотности, при низкой температуре. Например, ясень обыкновенный при температуре почвы 0 °C поглощает воды в 3 раза меньше, чем при +(20–30) °С. Способность поглощать воду при той или иной температуре зависит от приспособленности растений к тепловому режиму почв в местах их произрастания. Виды с ранним началом развития, как правило, могут всасывать воду корнями при более низкой температуре, чем развивающиеся позднее. Тундровые растения и некоторые деревья, растущие на почвах с подстилающей их многолетней мерзлотой, могут поглощать воду при температуре почвы 0 °C.
У высших растений есть и дополнительные пути поступления воды в тело. Мхи могут поглощать воду всей поверхностью, как и лишайники. Особенно много воды впитывают такие мхи, как кукушкин лен, виды сфагнума, чему способствует строение их листьев и побегов. При полном насыщении сфагновые мхи содержат в своем теле в десятки раз больше воды, чем в воздушно-сухом состоянии (рис. 30). Семена поглощают воду из почвы. Из воздуха, насыщенного водяными парами, в дождевом тропическом лесу поглощают воду многие эпифиты, например папоротник гименофиллум – тонкими листьями, многие орхидеи – воздушными корнями. В чашевидных влагалищах листьев многих зонтичных скапливается вода, которая постепенно всасывается эпидермисом. Виды из рода тилляндсия (бромелиевые) существуют в пустыне Атакама практически исключительно за счет влаги туманов и росы, которую впитывают чешуевидные волоски на листьях.
Поступившая в растение вода транспортируется от клетки к клетке (ближний транспорт) и по ксилеме во все органы, где расходуется на жизненные процессы (дальний транспорт). В среднем 0,5 % воды идет на фотосинтез, а остальная – на восполнение потерь от испарения и поддержание тургора. Вода испаряется со всех поверхностей, как внутренних, так и наружных, соприкасающихся с воздухом. Различают устьичную, кутикулярную и перидермальную транспирацию.
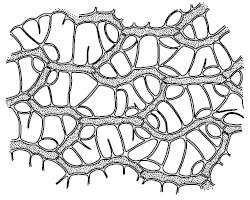
Рис. 30. Часть листа сфагнума болотного с клетками двух типов: зеленых с хлоропластами и бесцветных водоносных (Жизнь растений. Т. 4. 1978)
Через устьица транспирируется влага, испарившаяся с поверхности клеток внутри органов. Это основной путь расходования воды растением. Кутикулярная транспирация составляет менее 10 % от свободного испарения; у вечнозеленых хвойных пород она сокращается до 0,5 %, а у кактусов даже до 0,05 %. Относительно велика кутикулярная транспирация молодых развертывающихся листьев. Перидермальная транспирация обычно незначительна. Интенсивность общей транспирации повышается с увеличением освещенности, температуры, сухости воздуха и при ветре.
Водный баланс остается уравновешенным в том случае, если поглощение воды, ее проведение и расходование гармонично согласованы друг с другом. Нарушения его могут быть кратковременными или длительными. По приспособлениям наземных растений к кратковременным колебаниям условий водоснабжения и испарения различают пойкилогидрические и гомойогидрические виды.
У пойкилогидрических растений содержание воды в тканях непостоянно и сильно зависит от степени увлажнения окружающей среды. Они не могут регулировать транспирацию и легко и быстро теряют и поглощают воду, используя влагу росы, туманов, кратковременных дождей, в сухом состоянии находятся в анабиозе. Способны обитать там, где короткие периоды увлажнения чередуются с длительными периодами сухости.
Пойкилогидричность свойственна цианобактериям, всем водорослям, некоторым грибам, лишайникам (рис. 31), а также ряду высших растений: многим мхам, некоторым папоротникам и даже отдельным цветковым, по-видимому, вторично перешедшим к пойкилогидрическому образу жизни. Таков, например, южно-африканский кустарник Myrothamnus flabel-lifolia (розоцветные).

Рис. 31. Лишайник сосновых лесов Cetraria islandica – типичный пойкилогидрический организм (Жизнь растений. Т. 3. 1978)
В мелких клетках таллома большинства низших растений нет центральной вакуоли, поэтому при высыхании они равномерно сжимаются без необратимых изменений ультраструктуры протопласта. Сине-зеленые водоросли (цианобактерии), вегетирующие на поверхности почвы в пустыне, высыхая, превращаются в темную корочку. От редких дождей их слизистая масса набухает и нитчатые тела начинают вегетировать. Мхи, растущие на сухих скалах, стволах деревьев или на поверхности почвы лугов и степей (роды Thuidium, Tortula и др.), также могут сильно высыхать, не теряя жизнеспособности.
Пойкилогидричны пыльцевые зерна и зародыши в семенах растений.
Гомойогидрические растения способны поддерживать относительное постоянство обводненности тканей. К ним относят большинство высших наземных растений. Для них характерна крупная центральная вакуоль в клетках. Благодаря этому клетка всегда имеет запас воды и не так сильно зависит от изменчивых внешних условий. Кроме того, побеги покрыты с поверхности эпидермой с малопроницаемой для воды кутикулой, транспирация регулируется устьичным аппаратом, а хорошо развитая корневая система во время вегетации может непрерывно поглощать влагу из почвы. Однако способности растений, не выдерживающих высыхания, регулировать свой водный обмен различны. Среди них выделяют разные по экологии группы.
Дата добавления: 2015-11-26; просмотров: 1715;