МЕМБРАННЫЙ ПОТЕНЦИАЛ ДЕЙСТВИЯ
А. Характеристика ПД. ПД - электрический процесс, выражающийся в быстром колебании мембранного потенциала вследствие перемещения ионов в клетку и т клетки и способный распространяться без затухания(без декремента). Он обеспечивает передачу сигналов между нервными клетками, между нервными центрами и рабочими органами, в мышцах - процесс электромеханического сопряжения (рис. 3.3, а).
Величина ПД нейрона колеблется в пределах 80-110 мВ, длительность пика ПД нервного волокна составляет 0,5-1 мс. Амплитуда ПД не зависит от силы раздражения, она всегда максимальна для данной клетки в конкретных условиях: ПД подчиняется закону «все или ничего», но не подчиняется закону силовых отношений -закону силы. ПД либо совсем не возникает на раздражение клетки, если оно мало, либо он максимальной величины, если раздражение является пороговым или сверхпороговым. Следует отметить, что слабое (подпороговое) раздражение может вызвать локальный потенциал. Онподчиняется закону силы: с увеличением силы стимула величина его возрастает (подробнее см. раздел 3.6). В составе ПД различают три фазы: 1 фаза - деполяризация, т.е. исчезновение заряда клетки - уменьшение мембранного потенциала до нуля; 2 фаза - инверсия, изменение заряда клетки на обратный, когда внутренняя сторона мембраны клетки заряжается положительно, а внешняя - отрицательно (от лат. туегзю - переворачивание); 3 фаза - реполяризация, восстановление исходного заряда клетки, когда внутренняя поверхность клеточной мембраны снова заряжается отрицательно, а наружная - положительно.
Б. Механизм возникновения ПД.Если действие раздражителя на клеточную мембрану приводит к возникновению ПД, далее сам процесс развития ПД вызывают фазовые изменения проницаемости клеточной мембраны, что обеспечивает быстрое движение иона Ка+ в клетку, а иона К+ — из клетки. Величина мембранного потенциала при этом сначала уменьшается, а затем снова восстанавливается до исходного уровня. На экране осциллографа отмеченные изменения мембранного потенциала предстают в виде пикового потенциала - ПД. Он возникает вследствие накопленных и поддерживаемых ионными насосами градиентов концентраций ионов внутри и вне клетки, т.е. за счет потенциальной энергии в виде электрохимических градиентов разных ионов. Если заблокировать процесс выработки энергии, то ПД некоторый период времени будут возникать, но после исчезновения градиентов концентраций ионов (устранение потенциальной энергии) клетка генерировать ПД не будет. Рассмотрим фазы ПД.
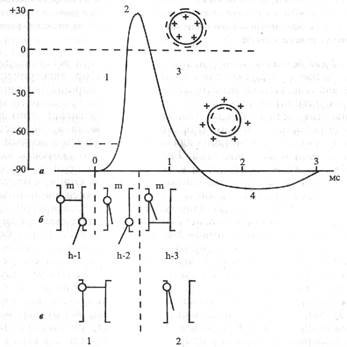
| Рис. 3.3. Схема, отражающая процесс возбуждения. а - потенциал действия, его фазы: 1 - деполяризация, 2 - инверсия (овершут), 3 - реполяризация, 4 — следовая гиперполяризация; б - натриевые ворота; (Ь-1 - в состоянии покоя клетки); в - калиевые ворота (1 - в состоянии покоя клетки). Знаки плюс (+) и минус (-) - знаки заряда внутри и вне клетки в различные фазы ПД. (См. пояснения в тексте.) Существует много различных названий фаз ПД (единого мнения не сложилось): 1) местное возбуждение - пик ПД - следовые потенциалы; 2) фаза нарастания - фаза спада -следовые потенциалы; 3) деполяризация - овершут (перехлест, превышение, перелет), причем эта фаза в свою очередь делится на две част: восходящая (инверсия, ОТ лат. шуегяю - переворачивание) н нисходящая (реверсия, от лат. геуегзю - возврат) - реполя-рнзапия. Имеются и другие названия. |
Отметим одно противоречие: термины «реполяризация» и «реверсия» но смыслу одинаковы - возврат к предыдущему состоянию, но эти состояния различны: в одном случае заряд исчезает (реверсия), в другом -восстанавливается (реполяршация). Наиболее корректны тс названия фаз ПД, в которых заложена общая идея, например изменение заряда клетки. В этой связи обоснованно использовать следующие названия фаз ПД: !) фаза деполяризации - процесс исчезновения заряда клетки до нуля; 2) фаза инверсии - изменение заряда клетки на противоположный. т. е. весь период ПД, когда внутри клетки заряд положительный, а снаружи - отрицательный; 3) фаза реполярпзацин - восстановление заряда клетки до исходной величины (возврат к потенциалу покоя).
1. Фаза деполяризации (см. рис. 3.3, а, 1). При действии деполяризующего раздражителя на клетку (медиатор, электрический ток) вначале уменьшение мембранного потенциала (частичная деполяризация) происходит без изменения проницаемости мембраны для ионов. Когда деполяризация достигает примерно 50% пороговой величины (порогового потенциала), возрастает проницаемость ее мембраны для иона Ка+, причем в первый момент сравнительно медленно. Естественно, что скорость входа ионов Ка* в клетку при этом невелика. В этот период, как и во время всей фазы деполяризации, движущей силой,обеспечивающей вход иона Na+ в клетку, являются концентрационный и электрический градиенты. Напомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются друг к другу), а концентрация ионов Na+ вне клетки в 10-12 раз больше, чем внутри клетки. При возбуждении нейрона повышается проницаемость его мембраны и для ионов Са+, но его ток в клетку значительно меньше, чем ионов Nа+. Условием, обеспечивающим вход иона Nа+ в клетку и последующий выход иона К* из клетки, является увеличение проницаемости клеточной мембраны, которая определяется состоянием воротного механизма ионных Nа- и К-каналов. Длительность пребывания электроуправляемого канала в открытом состоянии носит вероятностный характер и зависит от величины мембранного потенциала. Суммарный ток ионов в любой момент определяется числом открытых каналов клеточной мембраны. Воротный механизм ^-каналоврасположен на внешней стороне клеточной мембраны (Na+ движется внутрь клетки), воротный механизм К-каналов-на внутренней (К+ движется из клетки наружу).
Активация Nа- и К-каналов (открытие ворот) обеспечивается уменьшением мембранного потенциала, Когда деполяризация клетки достигает критической величины (Ekp, критический уровень деполяризации - КУД), которая обычно составляет -50 мВ (возможны и другие величины), проницаемость мембраны для ионов Nа+ резко возрастает - открывается большое число по-тенциалзависимых ворот Nа-каналов и ионы Nа+ лавиной устремляются в клетку. В результате интенсивного тока ионов Nа+ внутрь клетки далее процесс деполяризации проходит очень быстро. Развивающаяся деполяризация клеточной мембраны вызывает дополнительное увеличение ее проницаемости и, естественно, проводимости ионов Na+ - открываются все новые и новые активационные т-ворота Nа-каналов, что придает току ионов Na* в клетку характер регенеративного процесса.В итоге ПП исчезает, становится равным нулю. Фаза деполяризации на этом заканчивается.
2. Фаза инверсии. После исчезновения ПП вход Nа+ в клетку продолжается (m - ворота Na-каналов еще открыты - h-2), поэтому число положительных ионов в клетке превосходит число отрицательных, заряд внутри клетки становится положительным, снаружи - отрицательным. Процесс перезарядки мембраны представляет собой 2-ю фазу ПД - фазу инверсии (см. рис. 3.3, в, 2). Теперь электрический градиент препятствует входу Na+ внутрь клетки (положительные заряды отталкиваются друг от друга), проводимость Na* снижается. Тем не менее некоторый период (доли миллисекунды) ионы Na+ продолжают входить в клетку, об этом свидетельствует продолжающееся нарастание ПД. Это означает, что концентрационный градиент, обеспечивающий движение ионов Ка+ в клетку, сильнее электрического, препятствующего входу ионов Nа* в клетку. Во время деполяризации мембраны увеличивается проницаемость ее и для ионов Са2+, они также идут в клетку, но в нервных клетках роль ионов Са2+в развитии ПД мала. Таким образом, вся восходящая часть пика ПД обеспечивается в основном входом ионов Nа* в клетку.
Примерно через 0,5-1 мс после начала деполяризации рост ПД прекращается вследствие закрытия ворот Ка-каналов (Ь-3) и открытия ворот К-каналов (в, 2), т.е. увеличения проницаемости для ионов К+. Поскольку ионы К+ находятся преимущественно внутри клетки, они, согласно концентрационному градиенту, быстро выходят из клетки, вследствие чего в клетке уменьшается число положительно заряженных ионов. Заряд клетки начинает возвращаться к исходному уровню. В фазу инверсии выходу ионов К* из клетки способствует также электрический градиент. Ионы К* выталкиваются положительным зарядом из клетки ипритягиваются отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки - до конца фазы инверсии (см. рис. 3.3, а - пунктирная линия), когда начинается следующая фаза ПД - фаза реполяризации. Калий выходит из клетки не только по управляемым каналам, ворота которых открыты, но и по неуправляемым каналам утечки.
Амплитуда ПД складывается из величины ПП (мембранный потенциал покоящейся клетки) и величины фазы инверсии - около 20 мв.Если мембранный потенциал в состоянии покоя клетки мал, то амплитуда ПД этой клетки будет небольшой.
3. Фаза реполяризации. В этой фазе проницаемость клеточной мембраны для ионов К+ все еще высока, ионы К+ продолжают быстро выходить из клетки согласно концентрационному градиенту. Клетка снова внутри имеет отрицательный заряд, а снаружи - положительный (см. рис. 3.3, а, 3), поэтому электрический градиент препятствует выходу К* из клетки, что снижает его проводимость, хотя он продолжает выходить. Это объясняется тем, что действие концентрационного градиента выражено значительно сильнее действия электрического градиента. Таким образом, вся нисходящая часть пика ПД обусловлена выходом иона К+ из клетки. Нередко в конце ПД наблюдается замедление реполяризации, что объясняется уменьшением проницаемости клеточной мембраны для ионов К+ и замедлением выхода их из клетки вследствие закрытия ворот К-каналов. Другая причина замедления тока ионов К+ связана с возрастанием положительного потенциала наружной поверхности клетки и формированием противоположно направленного электрического градиента.
Главную роль в возникновении ПД играет ионNa*, входящий в клетку при повышении проницаемости клеточной мембраны и обеспечивающий всю восходящую часть пика ПД. При замене иона Nа+ в среде на другой ион, например холин, или в случае блокировки Na-каналов тетродотоксином, ПД в нервной клетке не возникает. Однако проницаемость мембраны для иона К+ тоже играет важную роль. Если повышение проницаемости для иона К+ предотвратить тетраэтиламмонием, то мембрана после ее деполяризации реполяризуется гораздо медленнее, только за счет медленных неуправляемых каналов (каналы утечки ионов), через которые К+ будет выходить из клетки.
Роль ионовСа2+ в возникновении ПД в нервных клетках незначительна, в некоторых нейронах она существенна, например в дендритах клеток Пуркинье мозжечка.
В. Следовые явления в процессе возбуждения клетки.Эти явления выражаются в гиперполяризации или частичной деполяризации клетки после возвращения мембранного потенциала к исходной величине (рис. 3.4).
Следовая гиперполяризацияклеточной мембраны обычно является следствием еще сохраняющейся повышенной проницаемости клеточной мембраны для К+. Ворота К-каналов еще не полностью закрыты, поэтому К + продолжает выходить из клетки согласно концентрационному градиенту, что и ведет к гиперполяризации клеточной мембраны. Постепенно проницаемость клеточной мембраны возвращается к исходной (натриевые и калиевые ворота возвращаются в исходное состояние), а мембранный потенциал становится таким же, каким он был до возбуждения клетки. Ионные помпы непосредственно за фазы потенциала действия не отвечают,ионы перемещаются с огромной скоростью согласно концентрационному и частично электрическому градиентам.
Следовая деполяризациятакже характерна для нейронов. Механизм ее изучен недостаточно. Возможно, она обусловлена кратковременным повышением проницаемости клеточной мембраны для Ка* и входом его в клетку согласно концентрационному и электрическому градиентам.
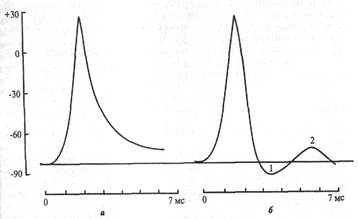
| Рис. 3.4. ПД двух клеток. а - замедление фазы реполяризации; б - следовые явления: 1 - следовая гиперполяризация; 2 - следовая деполяризация 3.5. ИССЛЕДОВАНИЕ ИОННЫХ ТОКОВ. ЗАПАС ИОНОВ В КЛЕТКЕ ПД обусловлен циклическим процессом входа №+ в клетку (восходящая часть пика) и последующим выходом К+ из клетки (нисходящая часть ПД), что является в свою очередь следствием активации и инактивации Na- и К-каналов. |
Наиболее растпространенный метод изучения функций ионных каналов - метод фиксации напряжения (voltage-clamp). Мембранный потенциал с помощью подачи электрического напряжения изменяют и фиксируют на определенном уровне, затем клеточную мембрану градуально деполяризуют, что ведет к открытию ионных каналов и возникновению ионного тока, который мог бы деполяризовать клетку. При этом пропускают электрический ток, равный по величине, но противоположный по знаку ионному току, поэтому трансмембранная разность потенциалов не изменяется. Это позволяет изучить величину ионного тока через мембрану. Применение различных блокаторов ионных каналов дает дополнительную возможность более глубоко изучить свойства каналов.
Количественное соотношение между ионными токами по отдельным каналам в покоящейся клетке и во время ПД и их кинетику можно выяснить с помощью метода локальной фиксации потенциала (patch-clamp). К мембране подводят микроэлектрод - присоску (внутри его создается разрежение) и, если на этом участке оказывается канал, исследуют ионный ток через него. В остальном методика подобна предыдущей. И в этом случае применяют специфические блокаторы каналов. В частности, при подаче на мембрану фиксированного деполяризующего потенциала было установлено, что через Ка-каналы может проходить и ион К+, но его ток в 10-12 раз меньше, а через К-каналы может проходить ион Ма+, его ток в 100 раз меньше, чем ток ионов К+.
Запас ионов в клетке, обеспечивающий возникновение возбуждения (ПД), огромен. Концентрационные градиенты ионов в результате одного цикла возбуждения практически не изменяются. Клетка может возбуждаться до 5 * 105 раз без подзарядки, т.е. без работы Ма/К-насоса. Число импульсов, которое генерирует и проводит нервное волокно, зависит от его толщины, что определяет запас ионов. Чем толще нервное волокно, тем больше запас ионов, тем больше импульсов оно может генерировать (от нескольких сотен до миллиона) без участия Nа/К-насоса. Однако в тонких волокнах на возникновение одного ПД расходуется около 1% концентрационных градиентов ионов Nа+и К*. Если заблокировать выработку энергии, то клетка будет еще многократно возбуждаться. В реальной действительности Nа/К-насос постоянно переносит ионы Nа+ из клетки, а ионы К+ возвращает в клетку, в результате чего поддерживается концентрационный градиент Nа+ и К+ за счет непосредственного расхода энергии, источником которой является АТФ. Имеются данные, что увеличение внутриклеточной концентрации Nа+ сопровождается повышением интенсивности работы Nа/К-насоса. Это может быть связано исключительно с тем, что для переносчика становится доступно большее количество внутриклеточных ионов Na+.
Дата добавления: 2015-10-22; просмотров: 2037;