Строение бактериальной клетки
Размеры — от 1 до 15 мкм. Основные формы: 1) кокки (шаровидные), 2) бациллы (палочковидные), 3) вибрионы (изогнутые в виде запятой), 4) спириллы и спирохеты (спирально закрученные).
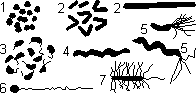
Формы бактерий:
1 — кокки; 2 — бациллы; 3 — вибрионы; 4—7 — спириллы и спирохеты.

Строение бактериальной клетки:
1 — цитоплазматическая мембрана; 2 — клеточная стенка; 3 — слизистая капсула; 4 — цитоплазма; 5 — хромосомная ДНК; 6 — рибосомы; 7 — мезосома; 8 — фотосинтетические мембраны; 9 — включения; 10 — жгутики; 11 — пили.
Бактериальная клетка ограничена оболочкой. Внутренний слой оболочки представлен цитоплазматической мембраной (1), над которой находится клеточная стенка (2); над клеточной стенкой у многих бактерий — слизистая капсула (3). Строение и функции цитоплазматической мембраны эукариотической и прокариотической клеток не отличаются. Мембрана может образовывать складки, называемые мезосомами (7). Они могут иметь разную форму (мешковидные, трубчатые, пластинчатые и др.).
На поверхности мезосом располагаются ферменты. Клеточная стенка толстая, плотная, жесткая, состоит из муреина (главный компонент) и других органических веществ. Муреин представляет собой правильную сеть из параллельных полисахаридных цепей, сшитых друг с другом короткими белковыми цепочками. В зависимости от особенностей строения клеточной стенки бактерии подразделяются на грамположительные (окрашиваются по Граму) и грамотрицательные (не окрашиваются). У грамотрицательных бактерий стенка тоньше, устроена сложнее и над муреиновым слоем снаружи имеется слой липидов. Внутреннее пространство заполнено цитоплазмой (4).
Генетический материал представлен кольцевыми молекулами ДНК. Эти ДНК можно условно разделить на «хромосомные» и плазмидные. «Хромосомная» ДНК (5) — одна, прикреплена к мембране, содержит несколько тысяч генов, в отличие от хромосомных ДНК эукариот она не линейная, не связана с белками. Зона, в которой расположена эта ДНК, называется нуклеоидом. Плазмиды — внехромосомные генетические элементы. Представляют собой небольшие кольцевые ДНК, не связаны с белками, не прикреплены к мембране, содержат небольшое число генов. Количество плазмид может быть различным. Наиболее изучены плазмиды, несущие информацию об устойчивости к лекарственным препаратам (R-фактор), принимающие участие в половом процессе (F-фактор). Плазмида, способная объединяться с хромосомой, называется эписомой.
В бактериальной клетке отсутствуют все мембранные органоиды, характерные для эукариотической клетки (митохондрии, пластиды, ЭПС, аппарат Гольджи, лизосомы).
В цитоплазме бактерий находятся рибосомы 70S-типа (6) и включения (9). Как правило, рибосомы собраны в полисомы. Каждая рибосома состоит из малой (30S) и большой субъединиц (50S). Функция рибосом: сборка полипептидной цепочки. Включения могут быть представлены глыбками крахмала, гликогена, волютина, липидными каплями.
У многих бактерий имеются жгутики (10) и пили (фимбрии) (11). Жгутики не ограничены мембраной, имеют волнистую форму и состоят из сферических субъединиц белка флагеллина. Эти субъединицы расположены по спирали и образуют полый цилиндр диаметром 10–20 нм. Жгутик прокариот по своей структуре напоминает одну из микротрубочек эукариотического жгутика. Количество и расположение жгутиков может быть различным. Пили — прямые нитевидные структуры на поверхности бактерий. Они тоньше и короче жгутиков. Представляют собой короткие полые цилиндры из белка пилина. Пили служат для прикрепления бактерий к субстрату и друг к другу. Во время конъюгации образуются особые F-пили, по которым осуществляется передача генетического материала от одной бактериальной клетки к другой.
Спорообразование у бактерий — способ переживания неблагоприятных условий. Споры формируются обычно по одной внутри «материнской клетки» и называются эндоспорами. Споры обладают высокой устойчивостью к радиации, экстремальным температурам, высушиванию и другим факторам, вызывающим гибель вегетативных клеток.
Размножение. Бактерии размножаются бесполым способом — делением «материнской клетки» надвое. Перед делением происходит репликация ДНК.
Редко у бактерий наблюдается половой процесс, при котором происходит рекомбинация генетического материала. Следует подчеркнуть, что у бактерий никогда не образуются гаметы, не происходит слияние содержимого клеток, а имеет место передача ДНК от клетки-донора к клетке-реципиенту. Различают три способа передачи ДНК: конъюгация, трансформация, трансдукция.

Конъюгация — однонаправленный перенос F-плазмиды от клетки-донора в клетку-реципиента, контактирующих друг с другом. При этом бактерии соединяются друг с другом особыми F-пилями (F-фимбриями), по каналам которых фрагменты ДНК и переносятся. Конъюгацию можно разбить на следующие этапы: 1) раскручивание F-плазмиды, 2) проникновение одной из цепей F-плазмиды в клетку-реципиента через F-пилю, 3) синтез комплементарной цепи на матрице одноцепочечной ДНК (происходит как в клетке-доноре (F+), так и в клетке-реципиенте (F-)).
Трансформация — однонаправленный перенос фрагментов ДНК от клетки-донора к клетке-реципиенту, не контактирующих друг с другом. При этом клетка-донор или «выделяет» из себя небольшой фрагмент ДНК, или ДНК попадает в окружающую среду после гибели этой клетки. В любом случае ДНК активно поглощается клеткой-реципиентом и встраивается в собственную «хромосому».
Трансдукция — перенос фрагмента ДНК от клетки-донора к клетке-реципиенту с помощью бактериофагов.
Вирусы
Вирусы открыты в 1892 г. Д.И. Ивановским при изучении мозаичной болезни табака (пятнистость листьев). Вирусы — неклеточные формы жизни. Проявляют признаки, характерные для живых организмов, только во время паразитирования в клетках других организмов. Вирусы являются внутриклеточными паразитами, но, в отличие от других паразитов, они паразитируют на генетическом уровне (ультрапаразиты). Существует несколько точек зрения на происхождение вирусов: 1) вирусы возникли в результате дегенерации клеточных организмов; 2) вирусы можно рассматривать как группу «потерявшихся», вышедших из-под контроля клетки генов («осколок генома»); 3) вирусы произошли от клеточных органоидов и др.
Вирусы состоят из нуклеиновой кислоты (ДНК или РНК) и белков, образующих оболочку вокруг этой нуклеиновой кислоты, т.е. представляют собой нуклеопротеидный комплекс. В состав некоторых вирусов входят липиды и углеводы. Вирусы содержат всегда один тип нуклеиновой кислоты — либо ДНК, либо РНК. Причем каждая из нуклеиновых кислот может быть как одноцепочечной, так и двухцепочечной, как линейной, так и кольцевой.
Размеры вирусов — 10–300 нм. Форма вирусов: шаровидная, палочковидная, нитевидная, цилиндрическая и др.
Капсид — оболочка вируса, образована белковыми субъединицами, уложенными определенным образом. Капсид защищает нуклеиновую кислоту вируса от различных воздействий, обеспечивает осаждение вируса на поверхности клетки-хозяина. Суперкапсид характерен для сложноорганизованных вирусов (ВИЧ, вирусы гриппа, герпеса). Возникает во время выхода вируса из клетки-хозяина и представляет собой модифицированный участок ядерной или наружной цитоплазматической мембраны клетки-хозяина.
Если вирус находится внутри клетки-хозяина, то он существует в форме нуклеиновой кислоты. Если вирус находится вне клетки-хозяина, то он представляет собой нуклеопротеидный комплекс, и эта свободная форма существования называется вирионом. Вирусы обладают высокой специфичностью, т.е. они могут использовать для своей жизнедеятельности строго определенный круг хозяев.

Вирусы, паразитирующие в бактериальных клетках, называются бактериофагами. Бактериофаг состоит из головки, хвостика и хвостовых отростков, с помощью которых он осаждается на оболочке бактерий. В головке содержится ДНК или РНК. Фаг частично растворяет клеточную стенку и мембрану бактерии и за счет сократительной реакции хвостика «впрыскивает» свою нуклеиновую кислоту в ее клетку.
Только паразитируя в клетке-хозяине, вирус может репродуцироваться, воспроизводить себе подобных.
Дата добавления: 2015-10-19; просмотров: 1553;