Надкласс четвероногие 2 страница
Из органов чувств у рептилий (за исключением змей и некоторых других видов) лучше, чем у амфибий, развиты глаза: они более подвижны. Приспособление их к видению на разных расстояниях (у многих видов на довольно больших) обеспечивается не только, как у рыб и амфибий, перемещением хрусталика, но и, как у высших позвоночных, путем изменения его кривизны. Кроме век, имеется мигательная перепонка. У ряда видов есть теменной орган, связанный с промежуточным мозгом и способный к восприятию световых раздражении. Некоторые прогрессивные изменения произошли в органе слуха: немного увеличился нижний мешочек; слуховая косточка лучше передает колебания барабанной перепонки во внутреннее ухо; кроме овального окошечка, появилось второе окошечко – круглое, что увеличивает подвижность экзолимфы, окружающей лабиринт; барабанная перепонка расположена не на поверхности головы, а в небольшом углублении. Органы обоняния развиты довольно хорошо, внутренняя поверхность их увеличилась. Органы вкуса у большинства видов развиты слабо. В осязании большую роль играет язык.
Скелет почти полностью окостенел благодаря замене грубой волокнистой костной ткани, характерной для земноводных, тонкой волокнистой, его прочность возросла, опорная и защитная функции улучшились. Череп увеличился; вместимость мозгового отдела стала большей, чем у амфибий. Боковая поверхность черепа у древних рептилий была сплошной, позднее в ней появились височные ямы – одна или две, что уменьшило его массу и увеличило поверхность для прикрепления жевательных мышц. Нижние челюсти, как и у амфибий, подвешены к мозговому отделу черепа при помощи квадратной кости и тоже состоят из нескольких костей (зубной, сочленовной и др.).
Тело первого позвонка (атланта) приросло в виде зубца к телу второго позвонка (эпистрофея). Кольцевидный атлант получил возможность вращаться вокруг зубца эпистрофея, благодаря чему стали возможны и повороты головы, а это имело большое значение для добывания пищи (в основном других животных) и для защиты от врагов. Атлант соединен с затылочным отделом черепа, с помощью одного мыщелка, что также способствует увеличению подвижности головы. Позвоночный столб разделяется на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой. Позвонки грудного отдела соединяются с грудиной хорошо развитыми ребрами, вследствие чего образуется грудная клетка, отсутствовавшая у амфибий. От поясничных позвонков отходят укороченные ребра, не доходящие до грудины. Крестцовых позвонков два (у амфибий один). В хвостовом отделе (за исключением черепах) позвонков много.
Кости ног лучше развиты и они более прочные, чем у земноводных. Между костями запястья и предплюсны имеются подвижные суставы. Поэтому предплечье и проксимальный ряд запястья, а также голень и проксимальный ряд предплюсны функционируют как целостные образования. Особо нужно отметить усиление поясов конечностей: плечевого – благодаря его соединениям с грудиной и грудным отделом позвоночника, тазового – вследствие того, что он опирается на два крестцовых позвонка и обе его половины образуют два сращения – седалищное и лонное.
Мышечная системаподверглась дифференциации, развились новые группы мышц, от которых зависят повороты головы, движения возникшей у рептилий шеи, расширение и сужение грудной клетки и др. Значительно увеличилась мускулатура ног, развились сгибатели и разгибатели пальцев. Масса и сила всей системы возросли.
Сердце, как и у амфибий, трехкамерное, но в желудочке снизу появляется зачаток перегородки. У крокодилов она уже полная, и сердце у них становится четырехкамерным (два предсердия и два желудочка). Следовательно, у пресмыкающихся усиливается разделение венозного и артериального токов крови, но полной изоляции обоих токов еще нет даже у крокодилов, как это видно из нижеследующего описания. В левое предсердие поступает артериальная кровь – она идет из легких. В правое предсердие поступает венозная кровь – она собирается из всех частей тела. Далее кровь из обоих предсердий поступает в желудочек: в правую его половину – венозная, в левую – артериальная. Благодаря наличию перегородки, хотя и неполной, кровь в желудочке меньше смешивается, чем у земноводных. Артериального конуса нет, и главные артериальные стволы отходят непосредственно от сердца.
От правой стороны желудочка отходят легочные артерии (бывшая четвертая пара жаберных артерий), несущие в легкие венозную кровь. Бывшая третья пара жаберных сосудов атрофировалась. Вторая пара сосудов состоит из двух дуг аорты: правая дуга начинается от левой части желудочка и несет артериальную кровь, левая дуга начинается от правой части желудочка (но ближе к левой части желудочка) и несет смешанную кровь. От правой дуги аорты отходят сонные артерии (бывшая первая пара жаберных сосудов), идущие к голове. Они снабжают мозг, особенно чувствительный к недостатку кислорода, артериальной кровью. Обе дуги аорты соединяются в аорту, которая разветвляется на множество артерий. Таким образом организм пресмыкающегося в основном снабжается смешанной кровью, но она насыщена большим количеством кислорода, чем у земноводных, что способствует более интенсивному обмену веществ.
Органами дыхания у рептилий служат легкие, так как их кожа непригодна для газообмена; слизистая оболочка ротовой полости, играющая значительную роль в дыхании земноводных, используется для выполнения этой функции немногими видами пресмыкающихся. По этой причине и особенно в связи с возросшей активностью рептилий по сравнению с амфибиями строение легких и дыхательных путей усложнилось. Легкие представляют собой органы большей величины и глубже расположенные в грудной полости, чем у земноводных. Внутренняя поверхность их значительно возросла благодаря развитию на ней множества перекладин с более густой сетью кровеносных сосудов. Воздух поступает в легкие, пройдя через хоаны, глотку, гортань, длинную трахею, просвет которой поддерживается хрящевыми кольцами. Во время прохождения через трахею воздух очищается от пыли, температура его приближается к температуре тела. У амфибий трахея была в зачаточном состоянии. Задний конец трахеи разделяется на два бронха, в стенках которых тоже имеются хрящевые кольца. Осуществление вдоха и выдоха происходит более эффективно, чем у амфибий, благодаря развитию у рептилий грудной клетки, объем которой увеличивается при вдохе и уменьшается при выдохе. Такой же механизм сохранился у высших позвоночных – птиц и млекопитающих.
Выделительная система сильно изменилась в связи с усилением интенсивности обмена веществ рептилий, что привело к увеличению количества продуктов диссимиляции, и полным приспособлением их к наземному образу жизни, вследствие чего возникла необходимость более экономно использовать поступающую в организм воду. Поэтому у рептилий развились более сложные органы выделения, названные тазовыми, или вторичными почками (метанефрос). Все канальцы этих почек не имеют воронок и начинаются боуменовыми капсулами, окружающими мальпигиевы тельца с увеличенным количеством капилляров. Канальцы тазовых почек очень длинные и густо оплетены капиллярами. В боуменовых капсулах из просочившихся туда из мальпигиевых телец растворенных в воде продуктов диссимиляции и некоторых органических веществ (например, глюкозы) образуется первичная моча. Последняя во время прохождения через длинные почечные канальцы отдает обратно в кровь через оплетающие их стенки капилляры значительную часть воды и другие вещества, необходимые для организма. Образовавшаяся после этого вторичная моча выходит не через вольфовы каналы, а через заново развившиеся мочеточники в клоаку и накапливается в мочевом пузыре. Процессы образования первичной и вторичной мочи совершаются и в туловищных почках, но с возвратом меньшего количества воды в кровь. Первичноводные же позвоночные, в особенности земноводные, кожа которых голая, часто страдают от избытка воды, а не от недостатка ее. Следует также отметить, что у рептилий конечным продуктом разложения бел ков является мочевая кислота, на выведение которой из организма с мочой затрачивается мало воды. У земноводных и у ряда других первичноводных позвоночных разложение белков приводит к образованию мочевины, на удаление которой требуется много воды.
У эмбрионов рептилий есть зачаток головной почки и развитая туловищная почка, которая функционирует у взрослых особей (у ряда видов ящериц) до наступления половой зрелости. Эти факты подтверждают преемственность трех видов почек в течение эволюции позвоночных.
Пищеварительная система более дифференцирована, чем у земноводных. Зубы в основном сосредоточены на челюстях, количество их велико, но они однотипны и служат преимущественно для захвата добычи. Слюнные железы лучше развиты, чем у амфибий (у ряда видов выделения этих желез ядовиты). У рептилий начинается процесс образования твердого неба, который завершается у крокодилов. Благодаря образованию твердого неба ротовая полость отделяется от носоглоточной, что облегчает как дыхание, так и поглощение пищи. Хорошо развиты (особенно у хищников, поедающих позвоночных животных) глотка, пищевод и желудок. Печень (с желчным пузырем) и поджелудочная железа более совершенны, чем у земноводных. Тонкая кишка значительно удлинена, толстая кишка короткая и заканчивается клоакой. Современные пре смыкающиеся питаются преимущественно животными, число растительноядных видов незначительно.
В состав мужской половой системы рептилий входят два семенника округлой формы, лежащие по бокам поясничного отдела позвоночника, два придатка семенников, образовавшиеся из передних участков туловищных почек, два семя провода (вольфовы каналы) и совокупительные органы, развившиеся из стенки клоаки. Женская половая система состоит из двух яичников, сильно разрастающихся в период размножения, и двух яйцеводов (мюллеровых каналов), впадающих в клоаку. Оплодотворение только внутреннее. Семя вводится самцами в клоаку самки при помощи совокупительных органов. Созревшие яйцеклетки, содержащие много желтка, попадают в воронки яйцеводов. Соединение женских и мужских гамет происходит в верхней части яйцеводов, куда проникают после полового акта сперматозоиды. Зиготы, продвигаясь по яйцеводам, в средней части последних окружаются белковой оболочкой, а в задней покрываются скорлупой. У некоторых видов ящериц и змей наблюдается партеногенетическое размножение самок. Самцов у таких видов, как правило, нет.
Развитие прямое; личинок у рептилий нет. Оплодотворенные яйца пресмыкающиеся откладывают в почву, под камнями, листьями и т. д. У ряда видов развитие зигот совершается в половых путях самки и сразу после выхода развившихся яиц наружу из них вылупляются сформированные животные, т. е. для таких видов характерно яйцеживорождение. У немногих видов наблюдается настоящее живорождение: сосуды желточного мешка тесно соединяются с сосудами яйцевода и эмбрион получает значительную часть питательных веществ из организма матери. Яйцеживорождение чаще наблюдается у видов, распространенных в северных и горных районах. Очевидно, яйцеживорождение является защитой развивающихся зародышей от охлаждения и других неблагоприятных условий. Нет ни одного вида рептилий, у которого яйца развиваются в воде; даже у рептилий, проводящих большую часть жизни в воде (все крокодилы, многие черепахи, некоторые змеи), откладка яиц и их развитие совершаются только на суше. Морские змеи, никогда не покидающие воду, относятся к живородящим.
Яйца и зародыши рептилий обладают рядом приспособлений для развития на суше. Во-первых, яйца покрыты скорлупой и другими оболочками, которые защищают их от высыхания и повреждений. Во-вторых, у зародышей развиваются особые зародышевые оболочки (серозная, амнион и аллантоис), которые имеют большое значение для обеспечения эмбрионального развития животных на суше. Серозная оболочка и амнион развиваются из одной и той же двойной складки тела зародыша, происходящей из эктодермы и мезодермы. Эта складка появляется с двух противоположных сторон в виде зачатков, которые растут навстречу друг другу и смыкаются над зародышем. Наружная часть складки превращается в серозу (ее наружный слой эктодермальный, внутренний – мезодермальный), а внутренняя – в амнион (его наружный слой мезодермальный, внутренний –эктодермальный). Полость, образуемая амнионом, наполнена водянистой жидкостью, благодаря чему развитие происходит во влажной среде, и зародыш защищен от высыхания и последствий различных сотрясений, особенно опасных в условиях воздушной среды.
Происхождение. Первые рептилии, возникшие во второй половине каменноугольного периода, обитали во влажных биотопах суши и, вероятно, размножались и развивались в воде. К этому времени суша уже была заселена низшими наземными растениями, различными беспозвоночными, начиналось завоевание воздуха насекомыми. Рептилии, которые могли удаляться на большие расстояния, чем амфибии, от водоемов, имели в изобилии пищу и не встречали серьезных врагов. В конце каменноугольного периода и неоднократно в течение пермского периода климат становился более засушливым, что вызывало гибель многих амфибий, и, наоборот, содействовало прогрессивному развитию и более широкому распространению на суше рептилий. Спустя некоторое время они перешли к размножению и развитию вне водоемов, сначала в более влажных наземных биотопах, а потом в более сухих местах. В течение мезозойской эры они распространились по всему земному шару и возникло много различных групп. Наряду с хищниками было много растительноядных видов, появились рептилии, приспособленные к полету, и рептилии, обитающие в морях. Величина их тела была различна, но большинство были крупными животными, а некоторые имели гигантские размеры – до 20 м и более в длину. Климат на протяжении почти всей мезозойской эры был теплым, очень благоприятным для этих животных, не имеющих достаточно высокую постоянную температуру тела и теряющих свою активность при охлаждении окружающей среды.
В конце мезозойской эры на земном шаре происходили мощные горообразовательные процессы, сопровождающиеся резким изменением условий существования (температуры, влажности и др.). Это повлекло за собой массовое вымирание мезозойских рептилий, не выдержавших борьбу за существование в таких тяжелых для них условиях неорганической среды, осложненную соревнованием с млекопитающими и птицами. Тогда же возникли и стали многочисленными в течение кайнозойской эры новые группы пресмыкающихся – ящерицы и произошедшие от них змеи. Из старых же групп сохранились лишь один очень примитивный вид первоящеров, небольшое количество крокодилов и черепах. Современные рептилии распространены преимущественно в теплых странах, особенно в субтропических и тропических, в странах с умеренным климатом их мало, на далеком севере они отсутствуют.
Систематический обзор
В мировой фауне насчитывается около 6600 видов рептилий: первоящеров – 1, черепах – 230, крокодилов – 22, ящериц – около 3900, змей около 2500. По численности видов рептилии стоят среди наземных позвоночных на втором месте, уступая только птицам.
Подкласс первоящеры (Prosauria)
Единственный представитель этой очень древней группы – гаттерия – сохранился только на островах Новой Зеландии. Строение гаттерии характеризуется рядом примитивных признаков: двуяковогнутые позвонки с остатками хорды между ними, на сошнике имеются зубы, совокупительных органов нет и т. д. Длина тела до 60 см. После заселения Новой Зеландии людьми гаттерия была почти истреблена; сейчас от полного истребления ее защищает специальный закон об охране этого замечательного животного.
Подкласс черепахи (Chelonia)
Отличаются от остальных рептилии прежде всего панцирем, состоящим из спинного и брюшного костных щитов, покрытых снаружи роговыми пластинками. Зубов нет, их заменяют заостренные роговые края челюстей. Скелет и другие системы органов довольно сильно изменены. Некоторые черепахи достигают огромной величины и имеют в длину до 2 м при массе 500 – 600 кг. Человек истребил большое количество черепах, так как мясо и яйца многих из них съедобны, а щиты идут на разные изделия. Большинство черепах временно или постоянно живут в воде. Интересно, что у последних панцирь сильно облегчен, а конечности превратились в ласты для плавания. Питаются черепахи преимущественно различными животными (беспозвоночными, рыбами и др.), но некоторые виды растительноядны.
Подкласс крокодилы (Crocodilia)
Крупные хищные пресмыкающиеся (самые большие из них достигают 10 – 12 м), ведущие водный образ жизни. Организация крокодилов в некоторых отношениях более совершенна, чем других рептилий (лучше развита нервная система, сердце четырехкамерное, зубы сидят в ячейках, имеется развитое твердое нёбо и т. д.). В настоящее время их осталось небольшое число видов и они все включены в Красную книгу. Крокодилы питаются крупными водными беспозвоночными, рыбами и другими позвоночными, в том числе млекопитающими. Некоторые виды нападают на человека. Из кожи крокодилов изготавливают различные изделия.
Подкласс чешуйчатые (Squamata)
Отряд ящерицы (Lacertilia). Большинство ящериц –животные небольших размеров. Обитают в самых разнообразных условиях: в степях, пустынях, лесах, горах и т. д. Мелкие виды питаются насекомыми и другими беспозвоночными, более крупные (вараны и др.) нападают на позвоночных; немногие виды растительноядны. Ящерицы истребляют вредных насекомых. Некоторых ящериц используют в пищу, но в общем практическое значение рассматриваемого отряда невелико. Известна лишь одна ядовитая ящерица – ядозуб, живущая на юге Северной Америки.
Отряд змеи (Ophidia). Своеобразная группа позвоночных, произошедших от каких-то вымерших ящериц. Следует отметить, что среди современных ящериц имеются виды, у которых наблюдается редукция ног. Змеи отличаются от своих предков не только отсутствием ног, но и рядом других признаков. Тело очень вытянутое, количество ребер увеличилось (на них при ползании змеи опираются). У удавов сохранились рудименты таза и задних ног. Грудины у змей нет. Веки срослись и стали прозрачными, что защищает глаза от разных повреждений. Среднее ухо и барабанная перепонка редуцировались. Одни органы (почки, половые железы и др.) сильно удлинились, другие уменьшились или атрофировались (левое легкое и др.) Почти все змеи – хищники и лишь некоторые виды питаются насекомыми. Кости, образующие верхнюю челюсть, подвижно соединены между собой и с соседними костями, а левая и правая половины нижней челюсти соединены связкой. Поэтому змеи могут широко раскрывать рот и заглатывать добычу больших размеров; стенки пищеварительной трубки тоже способны к значительному растяжению. Среди рассматриваемого отряда довольно много ядовитых видов. Яд выделяется видоизмененными верхнегубными слюнными железами и попадает в тело жертвы через раны, нанесенные особыми удлиненными зубами, расположенными на верхней челюсти. Эффективным средством против змеиного яда являются специальные сыворотки.
В нашей стране мало ядовитых змей, среди которых наиболее распространена гадюка обыкновенная (в степях южной части европейской территории – гадюка степная), близкая к гадюке, очень ядовитая гюрза. На домашних млекопитающих яд змей действует нередко сильнее, чем на человека. Однако польза, приносимая змеями, вероятно, значительно превышает причиняемый ими вред, так как они истребляют большое количество вредных грызунов, а яд разных змей идет на приготовление ряда лекарственных препаратов, в связи с чем некоторые виды этих пресмыкающихся разводят в специальных питомниках (серпентариях).
КЛАСС ПТИЦЫ (AVES)
Основные вопросы:
1. Класс Птицы (Aves)
2. Общая характеристика
3. Систематический обзор
Птицы возникли в начале юрского периода (по новейшим данным – в триасе) от пресмыкающихся, организация которых к этому времени уже сильно усложнилась в результате длительного эволюционного развития. Это сделало возможным переход древесных видов сначала к планирующему способу передвижения, а впоследствии к полету. Приспособление к полету наложило отпечаток на строение птиц и их жизнедеятельность. Отдел зоологии, посвященный всестороннему изучению птиц, называется орнитологией.
Общая характеристика
Тело птиц состоит из округлой головы, туловища, начинающегося очень подвижной шеей, передних конечностей, превратившихся в крылья, и хорошо развитых ног. Форма тела – обтекаемая, лишенная выступающих частей, компактная; крылья у передвигающихся по земле птиц или находящихся в покое сложены на. спине. Поверхность распростертых во время полета крыльев значительно превышает поверхность туловища. Отмеченные особенности имеют большое значение для обеспечения передвижения птиц в воздухе, а также по земле. Хвост отсутствует.
Кожа тонкая, что имеет существенное значение для уменьшения массы тела птиц и обеспечения подвижности перьев. В ней нет, как и у современных рептилий, желез, клетки наружных слоев ее эпидермиса сильно ороговевают, вследствие чего резко уменьшается испарение воды, столь необходимой для интенсивного обмена веществ, присущего птицам. Соединительнотканный слой кожи подстилается рыхлой клетчаткой, в которой могут накапливаться запасы жира.
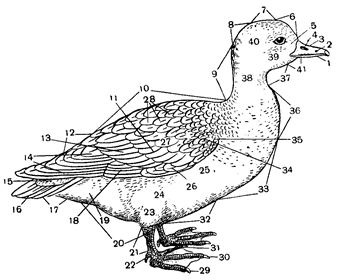
Рис. 68. Номенклатура отдельных частей тела и оперения птицы.
1 – подклювье; 2 – надклювье; 3 – клюв; 4 – ноздри; 5 – уздечка; 6 – лоб; 7 – темя; 8 – затылок; 9 – шея; 10 – спина; 11 – большие кроющие крыла; 12 – надхвостье; 13 – второстепенные маховые перья; 14 – первостепенные маховые перья; 15 – верхние кроющие хвоста; 16 – рулевые; 17 – нижние кроющие хвоста; 18 – кроющие маховых перьев; 19 – подхвостье; 20 – гузка; 21 – плюсна; 22 – задний палец; 23 – голень; 24 – бок брюха; 25 – бок груди; 26 – крылышко; 27 – средние кроющие крыла; 28 – плечевые; 29 – наружный палец; 30 – средний палец; 31 – внутренний палец; 32 – брюхо; 33 – грудь; 34 – малые кроющие крыла; 35 – кистевой сгиб крыла; 36 – зоб; 37 – шея; 38 – бок шеи; 39 – щеки; 40 – бока головы; 41 – подбородок.
Тело покрыто перьями. Различают перья контурные и пуховые (рис. 68). Контурное перо состоит из рогового стержня, который разделяется на очин и ствол; от ствола отходят длинные роговые пластинки – бородки первого порядка, от них – более короткие бородки второго порядка, сцепленные крючочками, в результате чего образуется прочная пластинка – опахало. В пуховом пере стержни слабые, бородки – мягкие, не сцепленные крючочками. Различают еще пух, в котором стержня нет, а имеются только мягкие бородки. Контурные перья прикреплены к коже не на всей поверхности, а только на определенных участках – птерилиях. Остальные участки кожи – аптерии – лишены контурных перьев, хотя они прикрыты ими. На этих участках у большинства птиц есть пух и пуховые перья. Последние могут быть и на птерилиях. Контурные перья названы так потому, что они в значительной степени определяют форму (контур) птиц. Они образуют прочную поверхность крыльев и на заднем конце тела – рулевую лопасть. Следовательно, контурные перья играют первостепенную роль в обеспечении полета. Весь перьевой покров предохраняет тело от излишней отдачи тепла и от механических повреждений, столь возможных во время полета, а также при передвижении по земле, на деревьях и т. д. Отмеченные положительные свойства перьевого покрова сочетаются с его малой массой, что так важно для летающих животных. Для сохранения эластичности контурных перьев птицы при помощи клюва смазывают их выделениями единственной кожной железы – копчиковой, расположенной около клоаки.
Образование пера начинается с выпячивания эктодермального сосочка, схожего с первыми стадиями развития роговых чешуи рептилий. Внутри сосочка имеется вырост соединительнотканното слоя кожи с кровеносными сосудами, питающими развивающееся перо. Снаружи сосочек окружен эпидермальным чехликом, под покровом которого происходит ороговение групп эктодермальных клеток и превращение их в бородки. По окончании развития бородок чехлик сбрасывается, освобождая готовое перо в виде пуха. При формировании пуховых и контурных перьев образуются стержни, от которых отходят бородки. Следовательно, готовые перья представляют собой чисто эктодермальные образования, в развитии которых принимает участие мезодермальный слой кожи.
Несмотря на достаточную прочность перьев, они все-таки повреждаются в результате воздействия разных внешних факторов, снашиваются (или, как говорят иногда, обнашиваются). Поэтому у птиц происходит периодическая смена оперения – линька. У одних птиц линьке одновременно подвергается весь перьевой покров. Такие птицы во время линьки не могут летать и прячутся в укромных местах. У других же птиц смена оперения совершается постепенно Линька первого типа наблюдается у видов, которые добывают пищу не в полете; линька второго типа характерна для видов, которые настигают свою жертву на лету. Смена перьев необходима также в связи с изменениями условий среды в разные времена года, например, в зимнем оперении больше пуха и пуховых перьев, чем в летнем и т. д.
Окраска перьев птиц разнообразна и характерна для каждого вида. Она обусловлена сочетанием разных пигментов: темных – меланинов, желтых и красных – липохромов. При отсутствии пигментов окраска перьев белая. Значение окраски различно: благодаря ей птицы менее заметны на фоне тех биотопов, где они обитают; она облегчает встречу самцов и самок одного и того же вида, общение между особями последнего, она меняется в зависимости от изменений неорганических условий среды, например, птицы, обитающие в более влажных и теплых местах их ареала, имеют более темную окраску, а в более сухих и холодных местах – более светлую. У ряда видов окраска может меняться в связи с изменением сезонных климатических условий.
Нервная система в связи с очень энергичным образом жизни птиц, потребовавшим усложнения всех систем органов, нервная система развита значительно лучше, чем у рептилий. Головной мозг намного превосходит по размерам мозг рептилий. Передний мозг велик, полосатые тела его, в которых находятся центры регуляции зрения, движения и других функций, весьма развиты. Кора полушарий выражена значительно лучше, чем у пресмыкающихся. В результате этих и других прогрессивных изменений роль переднего мозга как координатора всей сложной жизнедеятельности птиц резко возросла, способность к установлению новых связей со средой путем более быстрого приобретения условных рефлексов значительно улучшилась. Обонятельные доли развиты меньше, чем у ранее рассмотренных классов позвоночных, так как у птиц в их отношениях со средой главную роль играют зрение и слух. Усложнились и другие отделы головного мозга, в особенности мозжечок, имеющий очень крупные размеры. Значение его чрезвычайно возросло в связи с приспособлением птиц к полету. Спинной мозг ввиду редукции хвоста короткий, но строение его сильно усложнилось Головных нервов – 12 пар.
Глаза велики, их масса у многих видов превосходит массу головного мозга. Дальность зрения у многих видов очень велика, поле зрения весьма широко; аккомодация к видению на разных расстояниях совершенна и осуществляется разными способами – изменением кривизны хрусталика, а также роговицы, в меньшей степени более старым способом, т. е. перемещением хрусталика. Птицы хорошо различают цвета, их оттенки, особенности рисунка окраски окружающих их предметов. Необходимость столь совершенного зрения птиц вполне понятна, если учесть быстроту их передвижения во время полета, да и во многих случаях по земле.
Орган слуха устроен более сложно, чем у рептилий; нижний мешочек, воспринимающий звуковые колебания, увеличен за счет отходящего от него отростка (его называют завитком улитки, сравнивая с гомологичным органом млекопитающих, у которых этот отросток имеет 21/2 оборота); слуховая косточка благодаря изменениям в ее строении лучше передает в лабиринт звуковые колебания барабанной перепонки, расположенной в более развитом, чем у рептилий, наружном слуховом проходе; обрамление отверстия последнего перьями (особенно у совиных) напоминает слуховую раковину млекопитающих, играющую роль рупора. Птицы, как известно, издают разнообразные звуки, объединяющиеся у многих видов в сложные мелодии. В этом отношении птицы превосходят млекопитающих, способных издавать преимущественно однообразные звуки. Звуковая деятельность играет в жизни птиц огромную роль: в общении родителей с птенцами, самцов с самками, с другими особями того же вида, а также с представителями других видов.
Обоняние, как отмечалось, развито слабее, чем у других позвоночных, но у видов, питающихся падалью, которую они находят по запаху, оно развито лучше. Вкусовые раздражения воспринимаются особыми скоплениями клеток, расположенными на слизистой оболочке ротовой полости и языке. Но эти органы играют в жизни птиц несравненно меньшую роль, чем у млекопитающих. В коже имеются свободные окончания нервов и скопления разных чувствующих клеток, функции которых различны: осязание, восприятие изменений температуры, положения перьев и др.
Для нервной деятельности птиц характерны разнообразные и очень сложные инстинкты. Инстинктивная деятельность проявляется во взаимоотношениях самцов и самок, постройке гнезд, выкармливании и охране потомства, в способности многих птиц к дальним перелетам в связи со сменой времен года и т.д. Инстинкты проявляются на определенном этапе индивидуального развития птиц в определенных условиях внешней среды. Так, например, инстинкт ухаживания самцов за самками проявляется при достижении ими половой зрелости, наступление которой, в свою очередь, сильно зависит у ряда видов от удлинения светлого периода суток весной. Другие инстинкты свойственны птицам в течение всей жизни. Отсутствие сознания в инстинктивных действиях можно проиллюстрировать на следующем примере. Хорошо известно, что кукушки подбрасывают свои яйца в гнезда мелких воробьиных птиц. Вылупившийся кукушонок, еще слепой и глухой, выбрасывает яйца и птенцов хозяев гнезда, а те продолжают кормить быстро растущего, прожорливого нахлебника. Способность к образованию условных рефлексов у птиц развита во много раз лучше, чем у пресмыкающихся. Этой способностью люди пользуются при разведении домашних птиц. У кур, например, легко и быстро вырабатываются условные рефлексы: они сбегаются к месту кормления, вечером возвращаются на насесты и т. д.
Дата добавления: 2015-10-19; просмотров: 776;