Общая характеристика. Большинство видов этого класса приспособлено к быстрому плаванию, и форма их тела сходна с таковой акул
Большинство видов этого класса приспособлено к быстрому плаванию, и форма их тела сходна с таковой акул. Менее быстро плавающие рыбы имеют более высокое тело (например, у многих видов карповых рыб). Виды, ведущие малоподвижный образ жизни на дне (например, камбалы), имеют такую же уплощенную форму тела, как скаты (рис. 62).
Кожа подавляющего большинства костистых рыб покрыта небольшими костными, сравнительно тонкими чешуями, черепичеобразно налегающими друг на друга. Они хорошо защищают рыб от механических повреждений и обеспечивают достаточную гиб кость тела. Различают циклоидную чешую с закругленным верхним краем и ктеноидную с мелкими зубчиками на верхнем крае. Количество чешуи в продольных и поперечных рядах для каждого вида более или менее постоянное и учитывается при определении видовой принадлежности рыб. В холодное время рост рыб и чешуи замедляется или прекращается, поэтому на чешуе образуются годовые кольца, подсчитывая которые можно определить возраст рыбы. У ряда видов кожа голая, лишенная чешуи. В коже множество желез, выделяемая ими слизь уменьшает, как и у других рыбообразных, трение при плавании, защищает от бактерии и т. д. В нижних слоях эпидермиса имеются различные пигментные клетки, благодаря которым рыбы мало заметны на фоне окружающей их среды. У некоторых видов окраска тела может меняться в соответствии с изменениями окраски субстрата. Подобные изменения осуществляются пол влиянием нервных импульсов.
Величина тела рыб различна: от нескольких сантиметров до нескольких метров. В отличие от хрящевых и древних костных рыб среди костистых множество мелких видов, освоивших малые биотопы, недоступные для более крупных видов.
Размеры головного мозга по отношению к величине тела несколько больше, чем у хрящевых рыб. Передний мозг относительно мал по сравнению с другими отделами, но полосатые тела его велики и посредством связей их с другими отделами центральной нервной системы влияют на осуществление некоторых довольно сложных форм поведения; нервные клетки в его крыше отсутствуют. Промежуточный мозг и отделившиеся от него эпифиз и гипофиз хорошо развиты. Средний мозг крупнее других отделов головного мозга, в верхней части его две хорошо развитые зрительные доли. Мозжечок у хорошо плавающих рыб велик. Возросли размеры и усложнилось строение продолговатого и спинного мозга. Подчинение последнего головному мозгу по сравнению с тем, что наблюдается у хрящевых рыб, усилилось.
Некоторые усложнения произошли в органах чувств. Сейсмосенсорные органы расположены в каналах боковых стенок тела и образуют густую сеть на голове. Более развит круглый мешочек лабиринта, и вопреки долго существовавшему мнению, что костистые рыбы воспринимают лишь грубые сотрясения окружающей среды, многочисленными экспериментами доказано, что они реагируют на разнообразные звуковые колебания и используют их для общения внутри популяции. Способность к восприятию различных химических раздражении выражена очень хорошо. Многие виды воспринимают даже незначительные изменения температуры в окружающей их воде. Зрение костистых рыб рассчитано, как и у других рыб, на близкое расстояние; хрусталик шаровидный, не способный менять свою кривизну, резкость изображения достигается перемещением его при помощи сокращения особой мышцы –серповидного отростка.
Скелет. В течение эволюции рассматриваемого класса скелет постепенно окостенел; хорда сохранилась лишь у низших представителей класса, число которых незначительно. При изучении скелета нужно иметь в виду, что одни кости возникают в результате замещения хрящей костной тканью, другие развиваются в соединительнотканном слое кожи. Первые называются основными, вторые – покровными костями.

Рис. 63. Костистые рыбы. І – сельдь (сем. Сельдевые), ІІ – лосось (сем. Лососевые), ІІІ – карп (сем. Карповые), ІV – сом (сем. Сомовые), V – щука (сем. Щуковые), VІ – угорь (сем. Угревые), VІІ – судак (сем. Окуневые), VІІІ – бычок речной (сем. Бычковые), ІХ – камбала (сем. Камбаловые).
Мозговой отдел черепа представляет собой коробку, защищающую головной мозг и органы чувств: обоняния, зрения, равновесия и слуха. Крыша черепа образована парными носовыми, лобными, теменными костями. Последние примыкают к верхней затылочной кости, которая вместе с парными боковыми затылочными костями и основной затылочной костью образует заднюю часть черепа. Низ черепа состоит (спереди назад) из сошника, парасфеноида (широкой, длинной кости, очень характерной для черепа рыб) и основной кости. Передняя часть черепа занята капсулой, охраняющей органы обоняния; по бокам расположены кости, окружающие глаза, и ряд костей (обычно пять), защищающих органы слуха и равновесия.
Висцеральный отдел черепа состоит из ряда костных жаберных дуг, являющихся опорой и защитой жаберного аппарата и передней части пищеварительной системы. В состав каждой из упомянутых дуг входит несколько косточек. Дуг, к которым прикрепляются жабры, у большинства рыб пять (с каждой стороны). Внизу жаберные дуги соединяются между собой, а передняя из них связана с подъязычной дугой, которая состоит из нескольких костей. Верхняя из этих косточек – подъязычно-челюстная (гиомандибуляре) прикрепляется к мозговому отделу черепа в области слухового отдела и связана через квадратную кость с костями, окружающими ротовую полость. Следовательно, подъязычная дуга служит для соединения жаберных дуг с остальными частями висцерального отдела, а ее верхняя кость – с мозговым отделом черепа.
Края рта и вся ротовая полость укреплены рядом костей. Верхнечелюстной ряд костей представлен (с каждой стороны) межчелюстной и верхнечелюстной костями. Далее идет ряд костей: нёбная, несколько крыловидных и квадратная. Квадратная кость вверху примыкает к подвеску (гиомандибуляре), а внизу – к нижней челюсти. Последняя состоит из нескольких костей зубной (самой большой) угловой и сочленовной, соединяющейся с квадратной костью. У древних рыб (имевших еще хрящевой скелет) все дуги висцерального отдела черепа несли жабры, впоследствии же передние из этих дуг превратились в подъязычные дуги и челюстные ряды костей.
Позвоночный столб состоит из большого количества двояковогнутых (амфицельных) позвонков, в промежутках между которыми сохраняются остатки хорды. От каждого позвонка отходит вверх и несколько назад длинный остистый отросток. Основания этих отростков разделены, и они образуют канал, по которому проходит спинной мозг. От нижней стороны тел позвонков отходят два коротких поперечных отростка, к которым в туловищном отделе прикрепляются длинные изогнутые ребра. Они оканчиваются свободно в мышцах и образуют каркас боковых стенок тела. В хвостовой части тела от позвонков книзу отходят только нижние остистые отростки.
Органы передвижения у костных рыб, как и у хрящевых представлены непарным (спинные, анальный, хвостовой) и парным (грудные и брюшные) плавником. Лопасти плавников поддерживаются костными лучами. Одни лучи – мягкие, состоящие из ряда костных участков, другие – твердые, цельные, концы которых у многих видов заострены. Лучи спинных и анального плавников опираются на косточки – базалии (плавниковые подпорки), лежащие в мышцах у края тела. Парные плавники опираются на лежащие между мышцами пояса конечностей грудные – на плечевой (или передний) пояс, состоящий из нескольких костей верхняя из которых прикрепляется к черепу, брюшные – на тазовый (или задний) пояс, состоящий, как правило, из пары костей. Лучи хвостового плавника опираются на заднюю часть позвоночного столба. У низших костных рыб (как и у хрящевых) хвостовой плавник гетероцеркальный верхняя лопасть которого значительно больше нижней лопасти. У костистых рыб хвостовой плавник – гомоцеркальный, так как обе лопасти его приблизительно равны.
Постепенное окостенение скелета имело большое приспособительное значение в эволюции рыб т к оно способствовало развитию крепкой опоры для мышц и надежной защиты центральной нервной системы и внутренних органов. Кроме того, так как в состав костного скелета может входить большое количество мелких косточек то это явилось важным условием для возникновения множества видов, имеющих малую величину тела.
Костный скелет имеет большую массу, чем хрящевой, что могло бы затруднить плавание рассматриваемых рыб. Поэтому у последних развился путем ответвления от передней части кишки плавательный пузырь, лежащий над кишечником и заполненный смесью газов (азота, кислорода и углекислого газа), благодаря чему масса тела рыб значительно уменьшилась. У примитивных видов костных рыб (открытопузырных) пузырь остается соединенным с кишечной трубкой в течение всей жизни. У большинства видов (закрытопузырных}, появившихся позднее, он полностью отделяется от кишки. В стенках пузыря имеются густые сплетения капилляров, которые обеспечивают наполнение его газами. Объем газов в закрытом пространстве, как известно, может при сжатии сильно уменьшаться и, наоборот, при уменьшении сжатия резко увеличиваться. Увеличение и уменьшение объема пузыря происходит вследствие работы мышц, окружающих брюшную полость. Следовательно, плавательный пузырь не только уменьшает массу тела, но он выполняет также гидростатическую роль, т. е. облегчает подъем рыбы вверх при расширении его и погружение – при сжатии его. Плавательный пузырь редуцировался у ряда видов рыб, ведущих малоподвижный образ жизни на дне, и у тех видов, которые, обладая сильной мускулатурой, способны быстро перемещаться вверх или вглубь. Наличие пузыря у последних могло бы вызывать при быстром подъеме сильное расширение его и выворачивание внутренностей, как это было доказано многими наблюдениями.
У некоторых рыб плавательный пузырь, соединенный рядом косточек (веберов аппарат) с лабиринтом, способствует передаче некоторых звуковых волн в последний.
Сердце состоит из предсердия и желудочка, круг кровообращения один. Артериального конуса у костистых рыб нет, и артериальный сосуд, отходящий от желудочка, начинается луковицей аорты. Количество эритроцитов значительно больше, чем у хрящевых рыб, что способствует усилению интенсивности процессов диссимиляции. Селезенка хорошо развита.
Главными органами дыхательной системы являются жабры, состоящие из многих лепестков, прикрепленных проксимальными концами к жаберным дугам в отличие от жаберных пластин хрящевых рыб, прикрепленных одной стороной к межжаберным перегородкам. Следовательно, поверхность жабр костных рыб значительно больше, чем у хрящевых рыб. Более совершенен и механизм вдоха и выдоха. Довольно значительная часть газообмена (в среднем около 10 %) совершается через кожу. В газообмене могут принимать участие плавательный пузырь и некоторые части кишечника.
В выделительной системе особую роль играют туловищные почки, расположенные в виде двух длинных, темно-красных узких лент вдоль позвоночного столба. Дистальные концы мочеточников (вольфовых каналов) соединяются и открываются наружу общим отверстием, сзади анального отверстия. От дистальной части слившихся мочеточников отходит мочевой пузырь.
Пищеварительная системаимеет некоторые особенности. Спиральной складки нет, увеличение поверхности кишечника происходит за счет удлинения его: образуются петли. Кроме того, от начальной части кишки у многих высших костистых рыб отходят пилорические отростки, тоже увеличивающие поверхность кишки. Кал удаляется через анальное отверстие; клоаки нет. Строение рассматриваемой системы различно в зависимости от характера питания. У хищников, нападающих на других рыб и более крупных беспозвоночных, широкий рот, обычно усаженный большими, острыми зубами; желудок большой, резко отделенный от пищевода и начала кишки, общая длина кишечника значительно короче, чем у растительноядных видов. У последних и в особенности у видов, питающихся мелкими беспозвоночными и органическими остатками, зубы малы или их нет; желудок почти не выражен или отсутствует. У карповых и некоторых других рыб в глотке имеются особые глоточные зубы для механической обработки пищи. Печень хорошо развита, хотя не достигает такой величины, как у многих хрящевых рыб. Поджелудочная железа представлена отдельными дольками, находящимися в печени или в стенках начальной части кишечника, т. е. не имеет еще компактной формы, но развита лучше, чем у хрящевых рыб.
Изучение состава пищи разных рыб, особенно промысловых, имеет большое практическое значение, так как при наличии сведений о качественном и количественном составе органического мира определенных водоемов позволяет выяснить, какие виды и в каком количестве могут обитать в – каждом водоеме, при каком сочетании видов кормовая база водоемов может быть всесторонне использована без ущерба для последующего восстановления ее и т. д. Эти вопросы специально изучаются в курсах по рыбоводству.
Половая система костистых рыб по сравнению с хрящевыми и древними костными рыбами упрощена, но способна производить большое число гамет. Удлиненные два семенника (их часто называют «молоками») находятся под почками по бокам плавательного пузыря. Семя выходит не через вольфов канал, а через короткую трубку, образованную оболочкой семенника. Оба семяпровода открываются наружу общим половым отверстием сзади анального и мочевого отверстий или общим мочеполовым отверстием. Яичники (их обычно два) в период размножения представляют собой объемистые мешки, наполненные икрой, которая выходит наружу не через мюллеровы каналы (они редуцируются), а через короткие трубки, идущие от половых желез и открывающиеся наружу самостоятельным отверстием или в общее мочеполовое отверстие. Следовательно, созревшие яйцеклетки не попадают в полость тела, а быстро выходят из тела самок. Оплодотворение у подавляющего большинства видов костных рыб наружное. Внутреннее оплодотворение, а также живорождение свойственно сравнительно немногим представителям этого класса. Икра выметывается на водные растения и реже на другие подводные субстраты, а у меньшинства видов – в толщу воды. В связи с наружным оплодотворением масса гамет, зигот и мальков погибает. Поэтому плодовитость костных рыб очень велика. Например, карп производит более 1 млн. икринок, щука – около 1 млн., палтус – 2-3,5 млн., треска – до 1 млн. Созревание половых желез зависит от разных факторов – внутренних и наружных. Ускоряющее воздействие на созревание половых желез оказывают гормоны гипофиза (исследования Н. Л. Гербильского и др.).
Оплодотворение икры (как это было впервые доказано в 1857 г. В. П. Врасским) можно осуществить в искусственных условиях, смешивая икру и семя рыб в присутствии небольшого количества воды. В настоящее время такая методика хорошо разработана и успешно применяется в широких масштабах на так называемых рыбоводных заводах. Оплодотворенная икра развивается в благоприятных условиях обычно до стадии мальков, способных добывать пищу в естественных водоемах, куда их выпускают.
Такие мероприятия содействуют восстановлению ценных промысловых рыб, численность которых резко сократилась вследствие загрязнения водоемов, затруднения миграций рыб на места их размножения, усиленной добычи.
На скорость роста рыб влияют различные условия жизни: питание, температура, состав растворенных в воде веществ и др. Рост в зависимости от изменений условий жизни идет неравномерно. Известно, например, что он замедляется в холодное время, что можно проследить на годовых кольцах чешуи рыб. Исследования роста рыб в разных водоемах имеют большое теоретическое и практическое значение, так как благодаря им можно выяснить, какие условия благоприятствуют ему, какие, наоборот, задерживают его, как восстанавливаются запасы промысловых рыб и т. д.
Продолжительность жизни рыб различна: одни живут менее года (некоторые виды хамсы и др.), другие – несколько лет (тихоокеанские лососевые – кета, горбуша и др.), третьи – несколько десятков лет и даже около 100 лет (крупные осетровые, щука, сазан и др.).
Рыбообразные (бесчелюстные, рыбы) обладают приспособлениями (возникшими в течение длительной эволюции) для передвижения в воде, добывания пищи, спасения от хищников и т. п. На основе этих приспособлений у них развились закономерные передвижения, характерные для разных периодов их жизни – миграции. Самые короткие и частые из них суточные миграции. Зависят они от времени суток. Более длительные миграции – сезонные, зависят от сезона года. К ним, в частности, относят зимовальные миграции, когда рыбы уходят на глубину и в малоподвижном состоянии, не питаясь, пережидают неблагоприятное для них время года. Например, хамса, которая питается и размножается в Азовском море, зимует в Черном море, так как вода в нем не подвергается такому сильному охлаждению, как в мелководном Азовском море.
Длительные, у ряда видов далекие миграции, называемые кормовыми, совершают многие рыбы в места, где имеются в достаточном количестве организмы, которыми они питаются. Так, например, треска после окончания периода размножения в Атлантическом океане мигрирует в Баренцево море и некоторые другие северные моря.
Широко распространены у рыб нерестовые миграции, обеспечивающие размножение их иногда в очень далеких местах от районов, где они долго живут, питаются и растут. К таким видам относятся проходные рыбы, обитающие в морях, но размножающиеся в реках, впадающих в эти моря. Так, очень ценные осетровые рыбы (осетры, севрюга, белуга и др.) обитают в Каспийском, Азовском и Черном морях до наступления периода размножения, а для икрометания отправляются в верховья рек, впадающих в названные моря. После этого они возвращаются в моря, где живут до наступления следующего периода размножения. Мальки, развившиеся из отложенной и оплодотворенной в реках икры, сносятся течением в моря, где длительное время растут.
Такие рыбы размножаются несколько раз в течение жизни. Но есть рыбы, живущие несколько лет в океанах и морях, но размножающиеся только один раз в жизни. К ним относятся тихоокеанские лососи (кета, горбуша, чавыча и др.), которые для размножения идут в дальневосточные реки России, реки Северной Америки, Японии, Кореи, Китая, в которых происходит их нерест. Рыбы в результате погибают. Во время нерестовых миграций рыбы, как правило, не питаются. В отличие от упомянутых рыб пресноводный угорь, обитающий в течение длительного времени (до 20 и более лет) в реках Европы, никогда там не размножается, а совершает далекие миграции (7000 –8000 км) в Саргассово море, где нерестится и после этого погибает. Мальки угря подхватываются Гольфстримом и через два года попадают к берегам Европы и затем входят в реки.
Выделяют группу и полупроходных рыб, живущих в опресненных частях морей и размножающихся в нижнем течении рек, впадающих в эти моря. К ним относятся популяции воблы, сазана, леща, обитающие на севере Каспийского моря.
Миграции известны и у многих беспозвоночных животных, обладающих достаточно эффективными способами передвижения. У позвоночных они наиболее сложны и продолжительны вследствие высокого развития их нервной, двигательной и других систем органов.
Благодаря миграциям животные всесторонне используют различные части их ареалов. Миграции осуществляются наследственно закрепленными инстинктами, развившимися в результате длительного естественного отбора.
Исследования миграций имеют очень большое значение для охраны природы и целесообразного использования промысловых видов животных.
Систематический обзор
Ныне существующие костные рыбы принадлежат к двум подклассам: лучеперым и лопастеперым рыбам.
Подкласс лучеперые рыбы (Actinopterygn)
Парные плавники опираются на костные образования – радиалии, расположенные лучеобразно (рис. 153). Дыхание жаберное; хоан нет. Плавательный пузырь – непарный, лежащий над кишечником, возникший путем ответвления от него.
Надотряд ганоидные(Ganoidomorpha)
Эти рыбы были многочисленны в течение мезозойской эры. Тела их были защищены костными чешуями, покрытыми тонким слоем дентиноподобного вещества ганоина. Впоследствии, когда начался расцвет костистых рыб, подавляющее большинство их вымерло. Из четырех отрядов современных ганоидов здесь рассматривается один отряд, представители которого распространены в нашей стране.
Отряд осетрообразные. Большинство видов крупных размеров: длина тела белуги (рис. 63) до 4 –5 м, русского осетра – до 2 –3 м и т. д.; но стерлядь – небольшая рыба (длина тела – 40 –60 см). Хвостовой плавник – гетероцеркальный. На теле пять рядов костных пластинок (жучек), образовавшихся из ганоидных чешуи. Череп хрящевой, но окружен покровными костями. Хорда хорошо развита. Тел позвонков нет. Сохранилось брызгальце. Имеются артериальный конус и спиральная складка в кишке. Таким образом, у этих рыб, как и у других ганоидов, сохранился ряд признаков хрящевых рыб.
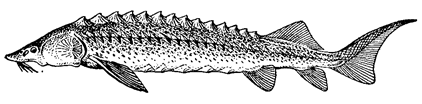
Рис. 64. Сибирский осетр.
Крупные осетровые – хищники, поедающие других рыб; остальные питаются мелкими беспозвоночными. Половозрелого состояния достигают поздно; самцы крупных видов в возрасте не ранее 10 –12, самки – 12 –15 лет; самцы стерляди – в 4 –5 лет, самки – 7 –8 лет. Большинство видов – проходные или полупроходные рыбы. Стерлядь обитает только в пресных водах, но совершает в реках довольно далекие нерестовые миграции. Всего известно 26 видов, из них 14 в России (главным образом в Каспийском и Азово-Черноморском бассейнах, в реках Сибири и Дальнего Востока). Издавна высоко ценились мясо и икра.
Надотряд костистые рыбы (Teleostei)
Тело покрыто костными чешуями, у некоторых видов вторично голое. Скелет в значительной степени или целиком костный. Остатки хорды сохраняются между телами позвонков. Вместо артериального конуса имеется луковица аорты. Спиральной складки в кишке нет, но у многих видов имеются пилорические отростки; кишечник удлинен.
В процессе эволюции костистых рыб происходили характерные, идущие в одном направлении изменения их организации: скелет постепенно окостеневал; в плавниках появились твердые заостренные лучи (у некоторых видов они превратились в колючки), брюшные плавники переместились вперед и расположились на уровне грудных плавников или впереди их; плавательный пузырь полностью отделился от передней части кишечника и наполнение его смесью газов происходит только через кровеносную систему; совершился ряд изменений в других органах, которые способствовали более быстрому плаванию, защите от врагов и увеличили общую приспособленность к обитанию в различных водоемах.
Практическое значение рыб общеизвестно. Они являются источниками белка и ряда других веществ, необходимых для полноценного питания людей. Население земного шара в течение XX в. и в особенности в последние десятилетия сильно увеличилось. В связи с этим добыча рыбы (а также водных моллюсков, ракообразных и других беспозвоночных) резко возросла. В дореволюционной России рыба добывалась в основном в реках, в других пресноводных водоемах и в южных морях страны.
В увеличении добычи рыбы значительную роль играет развитие прудового рыбоводства. Пруды – небольшие стоячие водоемы, большинство которых сооружают путем постройки плотин на небольших реках. Пруды наполняются водой за счет атмосферных осадков, подземных источников и рек.
В прудах рыба питается личинками насекомых (в основном личинками хирономид), рачками, червями и т. д. Для того чтобы увеличить рыбопродуктивность прудов, рыб подкармливают жмыхами, бобовыми растениями и др. Чаще всего в прудах выращивают различные расы карпа. Совместно с карпом можно в прудах разводить карасей, линей и другие виды рыб, не являющихся конкурентами в питании. В правильно организованных прудовых хозяйствах существует несколько типов водоемов (для размножения, выращивания мальков, нагула товарной рыбы, зимовки и т. д.). В таких хозяйствах можно получать с 1 га 15 –20 ц рыбы и более. Прудовое рыбоводство – одна из доходных отраслей животноводства. Необходимо так же интенсивно развивать рыбоводство во всех озерах и в других внутренних водоемах страны. В организации успешного прудового рыбоводства должны принимать участие, кроме рыбоводов, работники зоотехнии, ветеринарии и агрономии.
Эта огромная группа рыб разделяется на множество отрядов (30 – 40 и более). Здесь приводятся краткие сведения о некоторых из них в эволюционном порядке (т. е. от низших к высшим костистым рыбам).
Отряд сельдеобразные. Распространены во всех морях, чисто пресноводных мало; есть проходные виды. Многие имеют большое промысловое значение (разные виды сельдей, сардины, шпроты, хамса и др.).
Отряд лососеобразные. У многих видов на спине жировой плавник. Морские, проходные, некоторые чисто пресноводные. Промысловое значение очень велико (тихоокеанские лососи, семга, форель, нельма, омуль, хариус и др.).
Отряд щукообразные. В пресных водах северного полушария широко распространена обыкновенная щука, которую добывают в некоторых районах в большом количестве. Полезна как хищник, истребляющий малоценных сорных рыб.
Отряд угреобразные. Тело удлиненное, змееобразное, брюшных плавников нет. Подавляющее большинство видов обитает в морях, известен ряд проходных видов, в том числе европейский угорь, имеющий промысловое значение.
Отряд карпообразные. Пресноводные рыбы, число видов которых (свыше 2900) составляет 15 % всех видов класса. Многие имеют большое промысловое значение (сазан, лещ, карась, толстолобик, белый амур и др.). Выведены культурные породы сазана (разные породы карпа), карасей и др.
Отряд сомообразные. Обширный отряд, подавляющее большинство видов которого обитает в пресных водах, преимущественно в теплых. Тело, как правило, голое, около рта несколько пар усиков. Величина тела от гигантской (несколько метров) до карликовой (2 см). Многие хищники. Широко известен европейский сом, длина тела которого иногда достигает 5 м, а масса – 90-300 кг; хищник с широкой пастью, но мелкими зубами.
Отряд трескообразные. Большинство этих рыб имеют два или три спинных плавника. Брюшные плавники расположены в передней части брюха под грудными. Подавляющее большинство видов обитает в северных морях. Ряд видов принадлежит к числу важных промысловых объектов – треска, пикша, сайда, а в последнее время особенно хек, минтай, путассу. Лишь один вид проник из северных морей в пресные воды – налим, особенно многочисленный в Сибири. В печени ряда тресковых рыб накапливается много витамина D.
Отряд кефалеобразные. Брюшные плавники находятся посредине брюха, два спинных плавника. Распространены преимущественно в морских тропических и умеренных водах, в нашей стране главным образом в Черном море. Имеют важное промысловое значение.
Отряд окунеобразные. Самый большой отряд костистых рыб (свыше 6000 видов). Плавники с колючими лучами. Брюшные плавники находятся близко к переднему концу тела, под грудными плавниками или впереди их. Размеры тела от нескольких сантиметров до 4 – 5 м. Распространены на всем земном шаре в самых разнообразных водоемах. Ряд видов имеет большое промысловое значение: в пресноводных водоемах северного полушария – судак, окунь и др., в морях – ставрида, скумбрия, тунец, нототения и др.
Отряд камбалообразные. Хищные рыбы с уплощенным телом, лежащие во взрослом состоянии на дне на одном из боков. Брюшные плавники впереди грудных. Величина тела от 30 – 40 до 500 см. Мальки имеют обычное рыбообразное строение тела, приспособленное к плаванию в толще воды. Ряд видов имеет важное промысловое значение.
Подкласс лопастеперые рыбы (Sarcopterygii)
Эти рыбы по ряду признаков (кости, прикрывающие череп; жаберные крышки; костный скелет в парных плавниках; плавательный пузырь и др.) относятся к классу костных рыб. Но у них, как и у низших лучеперых рыб, сохранились примитивные признаки: развитая хорда, отсутствие тел позвонков, артериальный конус, спиральная складка в кишке и др. Сказанное свидетельствует о том, что лучеперые и лопастеперые рыбы произошли от древних костных рыб, живших в конце силура.
Однако пути эволюции обеих групп разошлись уже в начале девона. У лопастеперых рыб при сохранении указанных примитивных признаков сильно развились парные плавники с мощными опорными лопастями в основании их со скелетом из немногих подвижных удлиненных костей и появились парные плавательные пузыри, возникшие из задних жаберных мешков, лежащие на брюшной стороне, в которые поступал воздух через наружные ноздри и хоаны (внутренние ноздри). Эти древние лопастеперые рыбы обитали в небольших пресноводных водоемах и по разным причинам (высыхание воды, ухудшение качества ее, недостаток кислорода и кормовых объектов и т. п.) пытались выползать на берег, опираясь на плавники и используя для дыхания атмосферный воздух, и переходить в другие водоемы. Подкласс разделяется на два отряда: кистеперые рыбы и двоякодышащие рыбы.
Отряд кистеперые рыбы (Crossopterygii). Появились они в нижнем девоне. Обладали хорошо развитым опорным скелетом парных плавников. От них произошли в середине девона в результате длительного естественного отбора первые наземные позвоночные, дальнейшее приспособление которых к обитанию в прибрежных районах суши совершалось в каменноугольном периоде, и возникли примитивные земноводные – стегоцефалы. Последние широко распространились и начали вытеснять своих предков – кистеперых рыб.. Одна из групп последних в триасе переселилась в море, где они обитали до конца мезозойской эры. В отложениях кайнозойской эры остатки их не встречались, и возникло мнение, что морские виды кистеперых давно вымерли. Однако в 1938 г. в Индийском океане у берегов Южной Африки была обнаружена кистеперая рыба, близкая к давно вымершим морским мезозойским видам, названная латимерией. Впоследствии выяснилось, что эти рыбы постоянно обитают в районе Коморских островов (недалеко от Мадагаскара), где они изредка встречаются на довольно большой глубине. Длина их достигает 100 – 180 см; чешуя –крупная, массивная; парные плавники – мощные, массивные, очень подвижные; плавательный пузырь, т.е. бывшее легкое, для дыхания не приспособлен; хоан нет. Латимерии – хищные животные, питающиеся другими рыбами.
Отряд двоякодышащие рыбы(Dipnoi). Эти рыбы произошли от кистеперых рыб и были широко распространены с девона до пермского периода, позднее становились все более редкими. В настоящее время существует только шесть видов отряда один австралийский, четыре африканских и один южноамериканский. За исключением одного африканского вида, это крупные животные, достигающие в длину до 1,7 – 2 м. Они всеядные и даже хищные животные. Зубы у них срослись и превратились в массивные пластинки, способные дробить толстые раковины моллюсков. У австралийского вида одно легкое, у остальных – пара легких Кровь после окисления в легких идет в сердце, в котором появилась неполная перегородка, разделяющая предсердие на две части – правую и левую, как у земноводный. Они обитают в заболоченных, пересыхающих водоемах и могут довольно долго переносить отсутствие воды, окруженные грязевой камерой, используя для дыхания атмосферный воздух. Древние двоякодышащие рыбы, судя по их парным плавникам, могли переползать в другие водоемы. Но, вероятно, потомки кистеперых рыб оказались более приспособленными к освоению пограничных биотопов между водоемами и наземной средой и уже в девоне дали начало первым земноводным, с которыми двоякодышащие рыбы соревноваться не могли. Поэтому выжили те из них, которые могли длительное время пере носить в покоящемся состоянии отсутствие воды.
Дата добавления: 2015-10-19; просмотров: 1317;