Механизм шагательного рефлекса.
Рефлекс вызывается однократным раздражением кожи конечности. Он выражается в сгибании этой конечности с одновременным дополнительным разгибанием противоположной задней конечности – перекрестный разгибательный рефлекс. Затем согнутая конечность разгибается, опускается вниз, разогнутая – сгибается и поднимается вверх и т.д. Конечности опоры не касаются (собака фиксирована в станке с помощью лямок) и без дополнительного раздражения продолжают ритмично поочередно сгибаться и разгибаться – «шагать» в воздухе. Рефлекс может продолжаться часами, так как устранено влияние коры большого мозга. При неповрежденных сегментах, иннервирующих передние конечности (С5-Тh2), и более длительном раздражении задней конечности в рефлекс вовлекаются передние конечности. Это наблюдение показывает наличие межсегментарной связи на уровне спинного мозга, играющей важную роль в ритмических движениях конечностей. Поочередное сокращение и расслабление мышц-сгибателей и разгибателей осуществляется в результате взаимодействия процессов возбуждения и торможения в соответствующих центрах спинного мозга под влиянием импульсов, поступающих в мозг от проприорецепторов.
Особая роль проприорецепторов в осуществлении шагательного рефлекса определяется их расположением.
Мышечные веретена (мышечные рецепторы) расположены параллельно скелетной мышце: своими концами крепятся к соединительнотканной оболочке (перимизию) пучка экстрафузальных мышечных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5– 1 мм (рис. 10.2). Поэтому когда мышца расслабляется (удлиняется), растягиваются и мышечные рецепторы, что и ведет к их возбуждению (рис. 10.2, а).Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон (от лат. fusus – веретено), окруженных соединительнотканной капсулой. Длина интрафузальных мышечных волокон равна 4 – 7 мм, толщина – 15 –30 мкм. Длина экстрафузальных мышечных волокон – от нескольких миллиметров до многих сантиметров, толщина – 50 – 100 мкм. Вокруг средней части мышечного веретена обвивается несколько раз окончание одного афферентного волокна. Многие мышечные веретена иннервируются и другим (более тонким) афферентным волокном, которое также обвивает в виде спирали мышечный рецептор, но периферические его участки. Импульсы от мышечных рецепторов возбуждают нейроны своего центра (гомонимные нейроны) и тормозят нейроны центра-антагониста. Ближе к концам мышечного веретена подходят двигательные нервные окончания, являющиеся аксонами γ-мотонейронов спинного мозга. Их импульсация вызывает сокращение интрафузальных мышечных волокон (мышечных волокон рецептора), что ведет к возбуждению рецепторов.
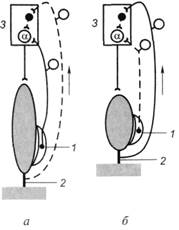
Рис. 10.2. Схема, отражающая процессы возбуждения a-мотонейронов спинного мозга и их торможения с помощью вставочных тормозных нейронов во время сокращения и расслабления скелетной мышцы:
а – мышца растянута и расслаблена – возбуждаются мышечные рецепторы (1);
б – мышца сокращена, напряжена и укорочена – возбуждаются сухожильные рецепторы (2); сплошные линии – импульсация выражена; пунктирные линии – импульсация отсутствует; 1– мышечные рецепторы (мышечные веретена); 2 – сухожилия мышцы и рецепторы (рецепторы Гольджи); 3 – полусегмент спинного мозга
Сухожильные рецепторы (рецепторы Гольджи) заключены в соединительнотканную капсулу и локализуются в сухожилиях скелетных мышц вблизи от сухожильно-мышечного соединения. Рецепторы представляют собой безмиелиновые окончания толстого миелинового афферентного волокна (подойдя к капсуле рецепторов Гольджи, это волокно теряет миелиновую оболочку и делится на несколько окончаний). Сухожильные рецепторы крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение при натяжении сухожилия (рис. 10.2, б).Поэтому сухожильные рецепторы посылают информацию в мозг о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы – о том, что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее).
Элемент шагательного рефлекса – поочередное сокращение и расслабление скелетной мышцы под влиянием импульсов от проприорецепторов, поступающих в свой центр.
Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим осмотонейронам спинного мозга и возбуждают их (рис. 10.2, а).Далее a-мотонейроны посылают импульсы к этой же скелетной мышце, что ведет к ее сокращению. Как только мышца сократилась (рис. 10.2, б),возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы. Импульсы от них поступают тоже сначала в свой центр в спинном мозге, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение a-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако расслабление мышцы (удлинение) ведет снова к возбуждению мышечных веретен и a-мотонейронов – мышца вновь сокращается. Вследствие сокращения мыщц возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что вновь ведет к расслаблению скелетной мышцы и т.д. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю. При этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу.
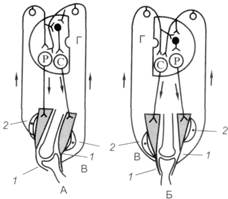
Рис. 10.3. Рефлекторные дуги шагательного рефлекса
А – конечность согнута; Б – контралатеральная конечность разогнута; В – коленные суставы с мышцами; Г – полусегменты спинного мозга; 1 – сухожилия и их рецепторы (рецепторы Гольджи); 2 – мышечные рецепторы (мышечные веретена); – афферентные пути от проприорецепторов; ¯ - эфферентные пути от центров сгибания (С) и разгибания (Р);
Для обеспечения поочередного сгибания и разгибания конечностей при шагательном рефлексе мышцы-сгибатели и мышцы-разгибатели должны сокращаться и расслабляться последовательно друг за другом. Причем если на одной ноге сокращены сгибатели, на другой ноге должны сокращаться разгибатели, что обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров сгибателей и разгибателей. На одноименной стороне при возбуждении центра мышцы-сгибателя тормозится центр мышцы-разгибателя (рис. 10.3).
Программы цепных двигательных актовшироко представлены в различных структурах мозга. Центральная нервная система хранит центральные программы как врожденных, так и выработанных действий. Так, например, дыхательные, глотательные и другие движения управляются врожденными моторными программами, тогда как различные спортивные, профессиональные навыки (акробатические упражнения, печатание на машинке и др.), т.е. быстрые точные движения, осуществляются по программам, формирующимся в течение жизни человека. При определенном опыте эти движения выполняются почти автоматически, в результате чего обратная афферентация, в том числе от проприорецепторов, перестает играть существенную роль (эту функцию выполняет мозжечок). Обратная афферентация становится нужной лишь при переделке, изменении навыка.
У позвоночных центральные программы запускаются управляющими центрами, а у беспозвоночных, как правило, командными нейронами, впервые выделенными в 1968 г. К. Вирсма. Однако сигнал от командного нейрона или управляющего центра играет лишь пусковую роль. Он не несет информации о том, как построить соответствующее движение. Структура движения заложена во взаимосвязях мотонейронов, с которыми он связан.
Ведущая роль в построении новых моторных программ принадлежит передним отделам коры больших полушарий (премо-торной, префронтальной коре).
В процессе индивидуальной жизни мозжечок обучается различным программам движения и сохраняет их. В этом процессе важную роль играют клетки Пуркинье. Мозжечок получает информацию о планируемом корой большого мозга движении, вносит коррекцию в план будущего движения и посылает ее в кору большого мозга. Быстрые мелкие точные движения осуществляются без сличения корой с импульсами от проприорецепторов. Мозжечок обучается также корригировать выполняемое движение, играет важную роль в регуляции тонуса мышц, поддержании позы, координации движения.
Базальные ганглии, представленные тремя парными образованиями (полосатое тело, включающее хвостатое ядро и скорлупу; бледный шар; ограда), имеют отношение к хранению программ врожденных двигательных актов и быстрых стереотипных движений. Врожденные программы имеются также в спинном мозге (шагательный рефлекс) и в стволе мозга (дыхания, глотания, выпрямительный рефлекс – восстановление нарушенной позы, ориентировочный рефлекс).
Инициация движенияпроисходит следующим образом. В моторной коре строится конечный и конкретный вариант моторного управления движением. Моторная кора использует оба принципа управления: контроль через петли обратной сенсорной связи и через механизм программирования. Это достигается тем, что к ней сходятся сигналы от мышечной активности, от сенсомоторной, зрительной и других отделов коры, которые и используются для моторного контроля коррекции движения. Кроме того, к моторной коре приходят сигналы, связанные с программированием движения из передних отделов коры и подкорки.
Моторная кора получает также информацию из структур, имеющих отношение к хранению двигательных программ. Так, в опытах с регистрацией нейронной активности у обезьян было показано, что при выполнении ими заученного движения активность нейронов зубчатого ядра мозжечка на 10 мс опережает изменение активности нейрона в моторной коре, которое предшествует появлению мышечного движения.
В моторную кору поступают также сигналы от базальных ганглиев – структура, которая ответственна за хранение главным образом двигательных программ врожденного поведения (пищевого, питьевого и др.), стереотипных движений. Клетки базальных ганглиев, так же как и мозжечка, разряжаются до начала движений, совершаемых животными в ответ на сигнал за 50 – 150 мс. В хвостатом ядре были найдены нейроны, у которых возбуждение опережает условную двигательную пищевую и оборонительную реакции. Эти реакции у нейронов появляются с выработкой условного рефлекса и исчезают с его утешением. Сигналы от базальных ганглиев и мозжечка распространяются не только вверх к коре, но и вниз, по нисходящему пути к спинному мозгу (так называемый экстрапирамидный путь).
В базальных ганглиях, мозжечке хранятся двигательные программы, которые могут быть реализованы с помощью сигнализации от специальных систем.
Согласно работам Г. Могенсона, все процессы управления движением включают три блока (рис. 10.4) и соответствующие им три фазы: 1) блок инициации движения, включающий лимбическую систему с прилегающим ядром (n. accumbens), и ассоциативную кору; 2) блок программирования движения, включающий мозжечок, базальные ганглии, моторную кору, таламус (как посредник между ними), а также спинальные и стволовые генераторы; 3) исполнительный блок, охватывающий мотонейроны и двигательные единицы. На рисунке (10.4) обратные связи опущены.
Произвольные движения человека – это сознательно регулируемые движения, которые осуществляются в отличие от непроизвольных с определенной целью.
У человека в отличие от животных цели произвольных движений формируются с помощью речевых процессов. Роль речевого опосредования в превращении непроизвольных импульсивных движений у детей в произвольные и сознательно управляемые была изучена А.Р. Лурия и его сотрудниками, которые показали, что активация детской речи – предварительное планирование в речевой форме предпринимаемых действий – ведет ребенка к овладению своим поведением, тогда как задержка в речевом развитии детей в патологических случаях приводит к снижению уровня произвольной (волевой) регуляции двигательного (поведенческого) акта. Произвольные движения, так же как и непроизвольные, являются результатом сопряженной работы лимбической системы и ассоциативной коры. Двигательная система работает как многоуровневая. На каждом уровне имеется своя «ведущая афферентация» и собственный тип регулируемых движений. Однако произвольные движения без пирамидной системы осуществляться не могут, а непроизвольные – могут. Непроизвольные движения запускаются посредством возбуждения экстеро- и проприорецепторов, могут осуществляться с помощью любого уровня ЦНС, в том числе и на уровне спинного мозга. Произвольные движения инициируются ассоциативной корой и лимбической системой с помощью механизмов памяти.
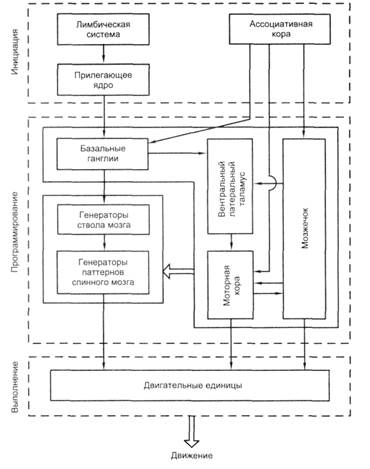
Рис. 10.4. Схема инициации двигательного акта (по Г. Могенсону, 1977, с изменениями)
Дата добавления: 2015-10-05; просмотров: 4448;