ЭВОЛЮЦИЯ ВИДОВ
Современные данные об эволюции животных указывают на расхождение линий эволюции и развитие параллельных линий (рис. 19.2 А), в том числе и среди млекопитающих (рис. 19.2 Б), и среди приматов (рис. 19.2 В). Это, в частности, означает, что сравнение строения мозга у грызунов, хищных и приматов или же у гориллы, шимпанзе и человека является исследованием не филогенетической, а адаптивной эволюции [Батуев, 1979], поскольку развитие шло в параллельных, независимых друг от друга линиях от общего, в каждом из ранее названных примеров своего, предшественника. Выводы о филогенетической эволюции видов и мозговых структур возможны лишь на основе анализа в последовательных рядах. Например, линия, ведущая к человеку, схематично может быть представлена следующим образом: рептилиеподобные млекопитающие – однопроходные – сумчатые – насекомоядные – обезьяны – приматы – большие обезьяны – африканские человекообразные обезьяны (рис. 19.2 Б, В), хотя и очевидно, что ныне живущие виды могут в той или иной степени отличаться от своих исходных форм – вымерших предков.
Прежде чем перейти к краткому описанию основных структурных различий мозга филогенетически разных животных и выяснению эволюционных закономерностей в развитии мозга, следует остановиться на вопросе о том, что является источником многообразия признаков, с которым имеет дело естественный отбор, и в результате каких «событий» появляются новые таксоны. Считается, что процессы развития находятся под генетическим контролем и что эволюцию следует рассматривать как результат изменений в генах, регулирующих онтогенез. Одним из оснований для заключения о том, что морфогенез, а тем самым и морфологическая эволюция (появление новых групп организмов) связаны не столько с изменениями структурных генов, сколько с изменениями регуляторных генов, управляющих генетической программой онтогенеза, являются данные, свидетельствующие о том, что морфологическая эволюция слабо коррелирует с молекулярной эволюцией (эволюцией белков). Так, например, лягушки за 150 млн. лет своей истории почти не изменились ни анатомически, ни по образу жизни, в то время как диапазон морфологического разнообразия у млекопитающих велик: от летучих мышей до китов, от слонов до человека. Несмотря на «морфологический консерватизм» лягушек, аминокислотные последовательности их белков подверглись значительным эволюционным изменениям. И, наоборот, человек и шимпанзе быстро дивергировали и многие систематики относят их к разным семействам, однако аминокислотные последовательности их белков на 99% одинаковы [Рэфф, Кофмен, 1986]. Другими аргументами в пользу морфогенеза за счет регуляторных генов служат данные об отсутствии корреляции между сложностью организации животного и величиной генома, наблюдаемые различия в величине генома у близкородственных животных, которые имеют сходную морфологическую сложность, а также то, что количество генов в геноме гораздо выше количества экспрессируемых (функционирующих) генов.
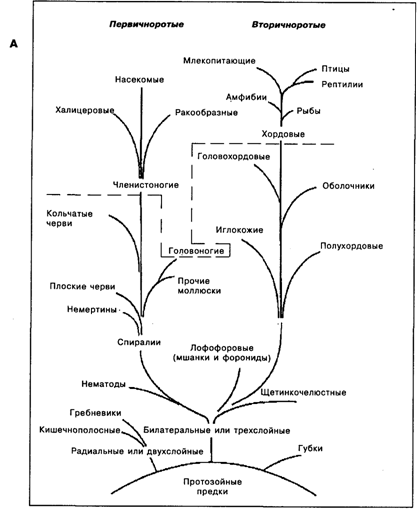
Рис. 19.2. А.Филогенетическое дерево Metazoa [по Рэфф, Кофман, 1986]. Пунктирная линия – условная граница между таксонами с элементарной сенсорной психикой и таксонами с перцептивной психикой [по Фабри, 1993]
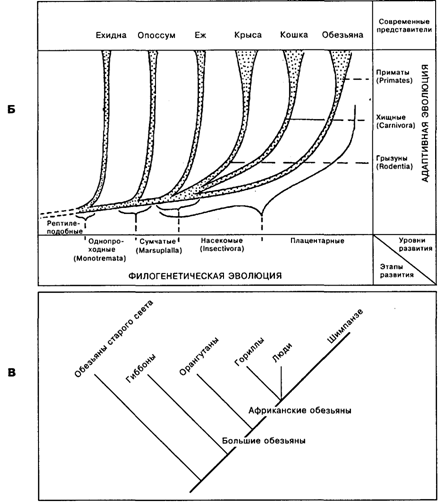
Б. Эволюционное дерево млекопитающих с выделением этапов филогенетической эволюции и уровней адаптивной эволюции [Батуев, 1979].
В. Филогенетическое дерево высших обезьян [Povinelli, Preuss, 1995]
Регуляторные гены являются «переключателями» между альтернативными путями развития, и количество их относительно невелико. Они функционируют на протяжении всего процесса развития, управляя процессом онтогенеза тремя различными способами: 1) регулируя время наступления тех или иных событий; 2) делая выбор из двух возможностей, тем самым определяя судьбу клеток и частей зародышей и 3) интегрируя экспрессию структурных генов и обеспечивая создание стабильных дифференцированных тканей. Поскольку регуляторный ген способен влиять на скорость роста какой-то определенной структуры, он контролирует как сроки появления, так и размеры данной структуры. А поскольку онтогенез слагается из связанных между собой процессов, т.е. формирование каждой отдельной структуры зависит как во времени, так и в пространстве от формирования других структур, изменения в сроках возникновения одного морфогенетического события могут иметь глубокие последствия, изменяя многие дальнейшие зависящие от него ступени онтогенеза [Рэфф, Кофмен, 1986]. Важно отметить, что модификации генетических регуляций приводят к возникновению новых структур, в том числе и разных «конструкций» мозга, а тем самым, и типов поведения, открывающих новые адаптивные возможности. Очевидно, что бифуркации на всех схемах, представленных на рис. 19.2, связаны именно с такими процессами появления новых признаков. В дальнейшем, после бифуркаций, изменения носят характер адаптивной эволюции, как это, например, показано на рис. 19.2 Б в ряду млекопитающих. Такой тип морфогенеза может объяснить «скачки» в развитии живого, связанные как с возникновением нервной системы и появлением психики, так и с формированием сложноорганизованного мозга человека с его отличной от других животных психикой, а также то, почему, даже приложив неимоверные усилия и создав все условия, не удастся «превратить» шимпанзе в человека, енота «заставить» говорить, а кролика – охотиться на слонов.
Наряду с зоологической, существует и психологическая классификация животных. Одной из них является классификация К.Э. Фабри, для создания которой он использовал представления А. Н. Леонтьева о «психике как форме отражения, позволяющей животному организму адекватно ориентировать свою активность по отношению к компонентам среды» [Фабри, 1993, с. 7], а в качестве уровней развития психики выделил уровень элементарной сенсорной психики и уровень перцептивной психики, граница между которыми проходит на уровне членистоногие, головоногие и хордовые (см. пунктирную линию на рис. 19.2 А), хотя оказалось, что не все крупные таксоны укладываются в эти рамки. Для высших позвоночных в этой классификации имеется стадия интеллекта, при том, что отмечаются принципиальные различия между интеллектом животных и людей, заключающиеся в том, что мышление животных всегда имеет конкретно-чувственный характер и осуществляется через действия с предметами. Для наиболее часто используемых определений психики (а именно, психика как психические процессы и состояния – субъективный мир – индивидуальный опыт) детальных психологических классификаций животного мира пока не создано.
Дата добавления: 2015-10-05; просмотров: 935;