Деятельность мобильных элементов
К мобильным элементам относят профаг mu, транспозоны, IS-элементы (Insertion Sequences), которые представляют собой дискретные сегменты ДНК, способные перемещаться внутри одного генома или между геномами, находящимися в одной клетке. Все мобильные элементы содержат гены, опосредующие процесс транспозиции, а также специфические инвертированные повторы на концах, которые тоже принимают участие в перемещении.
IS-элементы — это линейные фрагменты ДНК размером 0,2—2 т. п. н., содержащие на концах инвертированные повторы (фланкированные повторами). Существует несколько типов IS-элементов, отличающихся между собой последовательностью нуклеотидов, длиной, величиной инвертированных повторов (10—40 п. н.), частотой транспозиции (10-4 — 10-7 на поколение), а также длиной дуплицированных повторов в ДНК-мишени (5— 11 п. н.), которые образуются в процессе транспозиции. IS-элементы не содержат генов, определяющих фенотипически различимые свойства, а лишь информацию, необходимую для процесса транспозиции (структуру фермента транспозазы). При транспозиции этих элементов в новый сайт исходный IS-элемент остается на прежнем месте, т. е. инсерция сопровождается точным синтезом второй копии и не зависит от репликативных функций и рекомбинационного аппарата хозяина. Не требуется также никакой гомологии между IS-элементом и сайтом-мишенью. В разных репликонах содержится различное количество копий IS-элементов: например, нуклеоид E. coli содержит 4— 19 копий IS 1, 0— 12 копий IS2, 1—2 копии IS4.
Транспозоны (Tn) — это созданные на основе IS-элементов (или их производных) сложные мобильные структуры, содержащие гены, определяющие дополнительные (кроме транспозиции) функции. Все транспозоны фланкированы концевыми повторами. Часто ими являются известные IS-элементы, например IS 1, которые могут повторяться на концах транспозона в прямой или обратной (рис. 2.9) ориентации.
На рис. 2.9 видно, что с двух сторон центральная область транспозона (Km) фланкирована одинаковыми IS-элементами: IS-L (слева) и IS-R (справа), расположенными в Tn5 в разной ориентации. При этом сами IS-элементы также содержат инвертированные концевые повторы. Вся информация, необходимая для перемещения сложного транспозона, содержится в его IS-элементах: это та самая информация, которую используют сами IS-элементы при транспозиции, — гены, кодирующие транспозазу.
Разные мобильные элементы отличаются друг от друга степенью специфичности при выборе сайтов интеграции в репликоны. При высокой специфичности транспозон использует один или несколько сайтов ДНК-мишени, при низкой — множество предпочтительных либо практически любой сайт. Вероятность транспозиции зависит от свойств мобильного элемента, в первую очередь от его длины: при увеличении транспозона на 1 т. п. н. частота перемещения снижается в 2 раза. Частота транспозиции для одного и того же мобильного элемента тоже может быть неодинаковой, и зависит она от характера донорного и реципиентного репликонов. Так, известны мутации, которые могут подавлять частоту транспозиции. Кроме того, на перемещение мобильных элементов влияют факторы окружающей среды: температура, УФ-облучение, химические агенты.
Механизм транспозиции. Для объяснения механизма транспозиции, тонкости которого остаются невыясненными, предложено несколько моделей, принадлежащих к двум группам: репликативные и консервативные. Более понятной является репликативная (коинтегративная) модель. Согласно этой модели, на первой стадии происходит слияние молекул донорной и реципиентной ДНК, сопровождающееся дупликацией в местах слияния двух репликонов (рис.2.10).

На второй стадии репликоны разделяются благодаря реципрокной рекомбинации между идентичными участками (res-сайтами) в коинтеграте (рис. 2.10).
Для осуществления первой стадии необходима транспозаза и два инвертированных терминальных повтора. Считается, что транспозаза распознает именно эти участки и специфически взаимодействует с ними. Для разделения коинтеграта требуется другой продукт транспозоновых генов — резолваза, которая осуществляет сайт-специфическую рекомбинацию в res-сайтах.
При транспозиции путем коинтеграции принимают участие не только гены, принадлежащие самому мобильному элементу, но и репликативные функции клетки. Так, показано, что коинтеграты некоторых транспозонов могут диссоциировать только в RecA+ -клетках (где есть RecA-белок); иными словами, разрешение этих коинтегратов осуществляется при участии системы гомологичной рекомбинации.
Консервативная (простая, нерепликативная) модель транспозиции допускает выщепление (эксцизию) мобильного элемента из молекулы до-нора и вставку (инсерцию) его в молекулу-реципиент, т. е. не осуществляется дупликация самого мобильного элемента. Но и в этом случае соблюдается главная особенность процесса транспозиции: в сайте-мишени возникают дуплицированные повторы (рис. 2.11). Согласно консервативной модели, транспозаза катализирует ступенчатые двухнитевые разрывы реципиентной ДНК, аналогичные тем, что образуют рестриктазы. Транспозазы могут, вероятно, осуществлять надрезы на концах транспозона и соединять их с концами разрывов в ДНК-мишени. При этом фор-

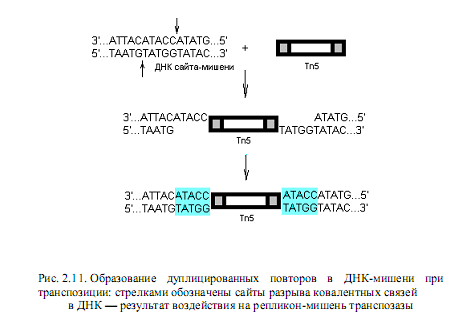
мируются однонитевые бреши, которые застраиваются репаративными системами клетки. В результате на концах мобильных элементов всегда создаются одинаковые короткие повторы участка ДНК-мишени (рис. 2.11). Некоторые мобильные элементы, например бактериофаг mu, могут участвовать в обоих типах транспозиции: в интегративной и консервативной.
Вырезание транспозонов из ДНК чаще всего осуществляется в результате гомологичной рекомбинации между копиями сайта-мишени.
Перемещения мобильных генетических элементов в пределах одного репликона могут приводить к образованию делеций и инверсий, а при межмолекулярной транспозиции могут возникать и другие мутации. Вообще, мобильные элементы индуцируют все виды хромосомных перестроек: слияние и диссоциацию репликонов, транслокации, делеции, инверсии, дупликации. Нередко встраивание мобильного элемента в регуляторный или кодирующий участок приводит к снижению экспрессии соответствующего гена. В некоторых случаях — наоборот: промотор, расположенный внутри самого транспозона, индуцирует экспрессию соседнего гена, который ранее был молчащим. Совместно с плазмидами и фагами мобильные элементы переносят гены между разными организмами и вносят весомый вклад в их изменчивость.
Дата добавления: 2015-09-29; просмотров: 1034;