Электрические синапсы.
Морфологически представляют собой слияние, или сближение, участков мембран. В последнем случае синаптическая щель не сплошная, а прерывается мостиками полного контакта. Эти мостики образуют повторяющуюся ячеистую структуру синапса, причем ячейки ограничены участками сближенных мембран, расстояние между которыми в синапсах млекопитающих 0,15—0,20 нм. В участках слияния мембран находятся каналы, через которые клетки могут обмениваться некоторыми продуктами. Кроме описанных ячеистых синапсов, среди электрических синапсов различают другие — в форме сплошной щели; площадь каждого из них достигает 1000 мкм, как, например, между нейронами ресничного ганглия.
Доля электрических синапсов (эфапсов) в ЦНС высокоорганизованных животных незначительна.
Пресинаптическая мембрана в таких синапсах сообщается с постсинаптической посредством миниатюрных мостиков, т.е. мембраны не разделены щелью. Потенциал действия, достигнув пресинаптической мембраны, преобразуется в градуальный ток, «стекающий» на постсинаптическую мембрану, где вновь возрождается в форме новой волны возбуждения. Таким путем возбуждение проводится через синапс почти без задержки.
Особенностью эфапсов является также их способность передавать возбуждение в двух противоположных направлениях, что принципиально невозможно в химических синапсах.
Синаптические контакты могут быть между аксоном и дендритом (аксодендритические), аксоном и сомой клетки (аксосоматические), аксонами (аксоаксональные), дендритами (дендродендритические), дендритами и сомой клетки.
Действие медиатора на постсинаптическую мембрану заключается в повышении ее проницаемости для ионов Na+. Возникновение потока ионов Na+ из синаптической щели через постсинаптическую мембрану ведет к ее деполяризации и вызывает генерацию возбуждающего постсинаптического потенциала (ВПСП) (см. рис. 2.19).
Для синапсов с химическим способом передачи возбуждения характерны синаптическая задержка проведения возбуждения, длящаяся около 0,5 мс, и развитие постсинаптического потенциала (ПСП) в ответ на пресинаптический импульс. Этот потенциал при возбуждении проявляется в деполяризации постсинаптической мембраны, а при торможении - в гиперполяризации ее, в результате чего развивается тормозной постсинаптический потенциал (ТПСП). При возбуждении проводимость постсинаптической мембраны увеличивается.
ВПСП возникает в нейронах при действии в синапсах ацетил холина, норадреналина, дофамина, серотонина, глутаминовой кислоты, вещества Р.
ТПСП возникает при действии в синапсах глицина, гамма-аминомасляной кислоты. ТПСП может развиваться и под действием медиаторов, вызывающих ВПСП, но в этих случаях медиатор вызывает переход постсинаптической мембраны в состояние гиперполяризации.
Существуют определенные различия в строении и функционировании возбуждающего и тормозного синапсов:
1. Синаптическая щель тормозного синапса уже, она составляет 20 нм (у возбуждающего - 30 нм).
2. Тормозные синапсы имеют более толстую и плотную постсинаптическую мембрану.
3. Пресинаптическая мембрана тормозного синапса содержит меньше пузырьков медиатора.
4. Медиатором в тормозных синапсах являются глицин, гамма - аминомасляная кислота (ГАМК), иногда - ацетилхолин.
Для включения в работу тормозного синапса опять нужен импульс возбуждения. При достижении импульсом пресинаптической мембраны в синаптическую щель выбрасывается квант тормозного медиатора. Действуя как химический раздражитель на мембранные рецепторы, тормозной медиатор изменяет проницаемость мембраны незначительно (в основном для ионов K+ и Cl-). Поскольку ионов K+ больше внутри клетки, а ионов Cl- в межклеточном пространстве, они начнут двигаться в противоположных направлениях: K+ - наружу, Cl- - внутрь. В результате этого увеличивается поляризованность мембраны (мембранный потенциал). При этом регистрируется положительное колебание с амплитудой около 5 mV - тормозной постсинаптический потенциал (ТПСП). Его продолжительность составляет (как и ВПСП) всего несколько миллисекунд. Но в течение этого времени никакое возбуждение на данном конкретном участке постсинаптической мембраны невозможно. В этом и заключается механизм действия тормозного синапса.
Для распространения возбуждения через химический синапс важно, что нервный импульс, идущий по пресинаптической части, полностью гасится в синаптической щели. Однако нервный импульс вызывает физиологические изменения в пресинаптической части мембраны. В результате у ее поверхности скапливаются синаптические пузырьки, изливающие медиатор в синаптическую щель.
Переход медиатора в синаптическую щель осуществляется путем экзоцитоза: пузырек с медиатором соприкасается и сливается с пресинаптической мембраной, затем открывается выход в синаптическую щель и в нее попадает медиатор. В покое медиатор попадает в синаптическую щель постоянно, но в малом количестве. Под влиянием пришедшего возбуждения количество медиатора резко возрастает. Затем медиатор перемещается к постсинаптической мембране, действует на специфические для него рецепторы и образует на мембране комплекс медиатор—рецептор. Данный комплекс изменяет проницаемость мембраны для ионов К+ и Na+, в результате чего изменяется ее потенциал покоя.
В зависимости от природы медиатора потенциал покоя мембраны может снижаться (деполяризация), что характерно для возбуждения, или повышаться (гиперполяризация), что типично для торможения. Величина ВПСП зависит от количества выделившегося медиатора и может составлять 0,12—5,0 мВ. Под влиянием ВПСП деполяризуются соседние с синапсом участки мембраны, затем деполяризация достигает аксонного холмика нейрона, где возникает возбуждение, распространяющееся на аксон.
В тормозных синапсах этот процесс развивается следующим образом: аксонное окончание синапса деполяризуется, что приводит к появлению слабых электрических токов, вызывающих мобилизацию и выделение в синаптическую щель специфического тормозного медиатора. Он изменяет ионную проницаемость постсинаптической мембраны таким образом, что в ней открываются поры диаметром около 0,5 нм. Эти поры не пропускают ионы Na+ (что вызвало бы деполяризацию мембраны), но пропускают ионы К+ из клетки наружу, в результате чего происходит гиперполяризация постсинаптической мембраны.
Такое изменение потенциала мембраны вызывает развитие ТПСП. Его появление связывают с выделением в синаптическую щель специфического медиатора. В синапсах разных нервных структур роль тормозного медиатора могут выполнять различные вещества. В ганглиях моллюсков роль тормозного медиатора выполняет ацетилхолин, в ЦНС высших животных — гамма-аминомасляная кислота, глицин.
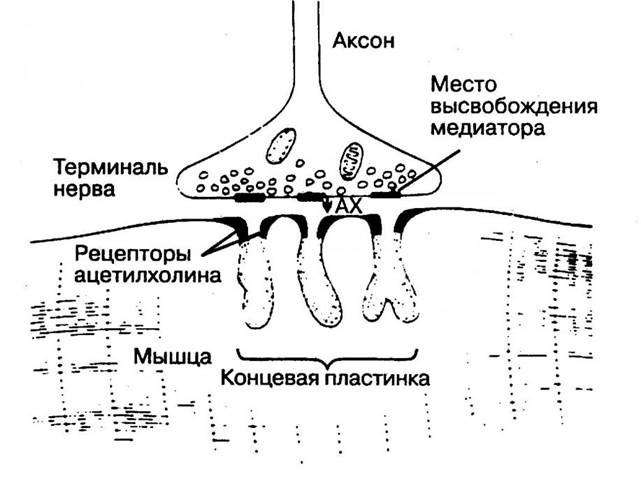
Нервно-мышечные синапсы обеспечивают проведение возбуждения с нервного волокна на мышечное благодаря медиатору ацетилхолину, который при возбуждении нервного окончания переходит в синаптическую щель и действует на концевую пластинку мышечного волокна. Следовательно, как и межнейронный синапс, нервно-мышечный синапс имеет пресинаптическую часть, принадлежащую нервному окончанию, синаптическую щель, постсинаптическую часть (концевая пластинка), принадлежащую мышечному волокну.
В пресинаптической терминали образуется и скапливается в виде пузырьков ацетилхолин. При возбуждении электрическим импульсом, идущим по аксону, пресинаптической части синапса ее мембрана становится проницаемой для ацетилхолина.
Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются ее кальциевые каналы. Ион Са2+ входит в пресинаптическую часть синапса из синаптической щели. Ацетилхолин высвобождается и проникает в синаптическую щель. Здесь он взаимодействует со своими рецепторами постсинаптической мембраны, принадлежащей мышечному волокну. Рецепторы, возбуждаясь, открывают белковый канал, встроенный в липидный слой мембраны. Через открытый канал внутрь мышечной клетки проникают ионы Na+, что приводит к деполяризации мембраны мышечной клетки, в результате развивается так называемый потенциал концевой пластинки (ПКП). Он вызывает генерацию потенциала действия мышечного волокна.
Нервно-мышечный синапс передает возбуждение в одном направлении: от нервного окончания к постсинаптической мембране мышечного волокна, что обусловлено наличием химического звена в механизме нервно-мышечной передачи.
Скорость проведения возбуждения через синапс намного меньше, чем по нервному волокну, так как здесь тратится время на активацию пресинаптической мембраны, переход через нее кальция, выделение ацетилхолина в синаптическую щель, деполяризацию постсинаптической мембраны, развитие ПКП.
Синаптическая передача возбуждения имеет ряд свойств:
1) наличие медиатора в пресинаптической части синапса;
2) относительная медиаторная специфичность синапса, т. е. каждый синапс имеет свой доминирующий медиатор;
3) переход постсинаптической мембраны под влиянием медиаторов в состояние де- или гиперполяризации;
4) возможность действия специфических блокирующих агентов на рецептирующие структуры постсинаптической мембраны;
5) увеличение длительности постсинаптического потенциала мембраны при подавлении действия ферментов, разрушающих синаптической медиатор;
6) развитие в постсинаптической мембране ПСП из миниатюрных потенциалов, обусловленных квантами медиатора;
7) зависимость длительности активной фазы действия медиатора в синапсе от свойств медиатора;
8) односторонность проведения возбуждения;
9) наличие хемочувствительных рецепторуправляемых каналов постсинаптической мембраны;
10) увеличение выделения квантов медиатора в синаптическую щель пропорционально частоте приходящих по аксону импульсов;
11) зависимость увеличения эффективности синаптической передачи от частоты использования синапса («эффект тренировки»);
12) утомляемость синапса, развивающаяся в результате длительного высокочастотного его стимулирования. В этом случае утомление может быть обусловлено истощением и несвоевременным синтезом медиатора в пресинаптической части синапса или глубокой, стойкой деполяризацией постсинаптической мембраны (пессимальное торможение).
Перечисленные свойства относятся к химическим синапсам. Электрические синапсы имеют некоторые особенности, а именно: малую задержку проведения возбуждения; возникновение деполяризации как в пре-, так и в постсинаптической частях синапса; наличие большей площади синаптической щели в электрическом синапсе, чем в химическом.
Синаптические медиаторы являются веществами, которые имеют специфические инактиваторы. Например, ацетилхолин инактивируется ацетилхолинэстеразой, норадреналин — моноаминоксидазой, катехолометилтрансферазой.
Неиспользованный медиатор и его фрагменты всасываются обратно в пресинаптическую часть синапса.
Ряд химических веществ крови и постсинаптической мембраны изменяет состояние синапса, делает его неактивным. Так, простагландины тормозят секрецию медиатора в синапсе. Другие вещества, называемые блокаторами хеморецепторных каналов, прекращают передачу в синапсах. Например, ботулинический токсин, марганец блокируют секрецию медиатора в нервно-мышечном синапсе, в тормозящих синапсах ЦНС. Тубокурарин, атропин, стрихнин, пенициллин, пикротоксин и др. блокируют рецепторы в синапсе, в результате чего медиатор, попав в синаптическую щель, не находит своего рецептора.
В то же время выделены вещества, которые блокируют системы, разрушающие медиаторы. К ним относят эзерин, фосфорорганические соединения.
В нервно-мышечном синапсе в норме ацетилхолин действует на синаптическую мембрану короткое время (1—2 мс), так как сразу же начинает разрушаться ацетилхолинэстеразой. В случаях, когда этого не происходит и ацетилхолин не разрушается на протяжении сотни миллисекунд, его действие на мембрану прекращается и мембрана не деполяризуется, а гиперполяризуется и возбуждение через этот синапс блокируется.
Блокада нервно-мышечной передачи может быть вызвана следующими способами:
1) действие местноанестезирующих веществ, которые блокируют возбуждение в пресинаптической части;
2) блокада высвобождения медиатора в пресинаптической части (например, ботулинический токсин);
3) нарушение синтеза медиатора, например при действии гемихолиния;
4) блокада рецепторов ацетилхолина, например при действии бунгаротоксина;
5) вытеснение ацетилхолина из рецепторов, например действие кураре;
6) инактивация постсинаптической мембраны сукцинилхолином, декаметонием и др.;
7) угнетение холинэстеразы, что приводит к длительному сохранению ацетилхолина и вызывает глубокую деполяризацию и инактивацию рецепторов синапсов. Такой эффект наблюдается при действии фосфорорганических соединений.
Специально для снижения тонуса мышц, особенно при операциях, используют блокаду нервно-мышечной передачи миорелаксантами; деполяризующие мышечные релаксанты действуют на рецепторы субсинаптической мембраны (сукцинилхолин и др.), недеполяризующие мышечные релаксанты, устраняющие действие ацетилхолина на мембрану по конкуренции (препараты группы кураре).
Мозг пользуется несколькими медиаторами. Норадреналин, дофамин, серотонин вызывают возбуждение, глицин и гамма - аминомасляная кислота (ГАМК) - торможение. Ацетилхолин - универсальный медиатор. Он может вызывать возбуждение и торможение. Каждый синапс для своей работы использует только один медиатор.
В мозговой ткани встречаются и смешанные синапсы. Такой синапс частично может проводить возбуждение как эфапс, а частично - с помощью медиатора.
Полагают, что некоторые лекарственные вещества, используемые при депрессиях и тревожности, воздействуют на химическую передачу в синапсах (Таблица 2).
Многие транквилизаторы и седативные средства (имипрамин, резерпин, др.) производят лечебный эффект, взаимодействуя с медиаторами, их рецепторами или отдельными ферментами. Галлюциногены типа диэтиламида лизерговой кислоты и москалина воспроизводят действие каких-то природных медиаторов мозга или подавляют действие других медиаторов. Сходный эффект с героином и морфием вызывают эндорфины, способные взаимодействовать с опиатными рецепторами. Из эндорфинов лучше всего изучена группа пептидов – энкефалинов (мет-энкефалин, β- эндорфин и др.)
Таблица 2
Дата добавления: 2015-09-28; просмотров: 2047;