Биоэлектрические потенциалы в возбудимых тканях.
Первые опыты по электрофизиологии были проведены в 1794 году. Итальянский ученый Л.Гальвани приготовил из лапки нервно-мышечный препарат (седалищный нерв с икроножной мышцей), надрезал мышцу и набрасывал нерв стеклянными крючками таким образом, чтобы часть его касалась поврежденного участка, и другая часть – неповрежденного. В момент набрасывания нерва мышца сокращалась. Как доказал позже Э.Дюбуа-Реймон, мышца сокращалась из-за электрического тока, источником которого была разность потенциалов между поврежденным и неповрежденным участком мышцы (поврежденный участок заряжен отрицательно, целый - положительно). Кстати, Э.Дюбуа-Реймон ввел в физиологию понятия «токи покоя» и «токи действия», которые в настоящее время называют потенциалом покоя и потенциалом действия.
К концу ХIХ и в начале ХХ стало известно, что электрические потенциалы образуются во всех живых тканях, а возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки.
До 50-х годов физиологи опирались на теорию происхождения биоэлектрических потенциалов В.Ю.Чаговца (1896), в основе которой была теория электролитической диссоциации. Суть теории: в раздражаемом или поврежденном участке увеличивается обмен веществ, образуется как продукт распада Н2СО3, диссоциирующая на Н+ и НСО3¯. Катионы Н+, по мнению В.Ю.Чаговца, диффундируют из возбужденного участка быстро, а анионы НСО3 – медленно, следствием чего и является возникновение разности потенциалов.
В 1902 году Ю.Бернштейном была предложена мембранная теория возникновения электрических потенциалов живых тканей. Суть ее – потенциалы возникают за счет разности зарядов внутри и снаружи клетки.
В настоящее время для исследования электрических явлений в клетки применяют микроэлектроды (стеклянные пипетки с тонким кончиком – 0,5 мкм), заполненные электролитом.
Было установлено, что величина внутриклеточного электрического потенциала у нервных клеток и волокон (например, у нервных гигантских клеток кальмара) в покое составляет 70 милливольт (мВ). Эту величину называют мембранным потенциалом покоя (МПП).
Природа поляризации клеточной мембраны сегодня в основном определена. Мембрана аксона кальмара, например, как и других клеток, тонкая, но достаточно прочная, Согласно данным электронной микроскопии ее толщина составляет 5-10 нм. Состоит мембрана из липидов, белков, мукополисахаридов (Рис.5). Мембрана легко проницаема для жирорастворимых веществ, молекулы которых легко проникают через липидный матрикс.
Крупные водорастворимые молекулы, в том числе анионы органических кислот практически не проникают мембрану и покидают клетку лишь путем экзоцитоза.
В то же время в мембране нервного волокна существуют каналы, проницаемые для воды, для малых молекул водорастворимых веществ и для малых ионов. Особое значение имеют каналы, проницаемые для ионов Nа +, К+, Cl -, Са ²+. В нервной мембране обнаружены специфические (селективные) натриевые, калиевые, хлорные и кальциевые каналы, то есть каналы избирательно пропускающие названные ионы. Эти каналы могут быть открытыми или закрытыми, то есть обладают воротными механизмами. В покое практически все натриевые каналы мембраны клетки закрыты, а большинство калиевых каналов открыты.
Для возникновения МПП, т.е. мембранного потенциала покоя, важно отличие ионного состава плазмы клетки от ионного состава внешней среды. В цитоплазме меньше ионов Nа+ и больше К+ и несравненно больше органических анионов. Катионные различия объясняются результатами работы так называемого калиево-натриевого насоса мембраны, непрерывно откачивающего Nа+ из клетки в обмен на К+. Движение ионов через мембрану называется активным ионным транспортом.

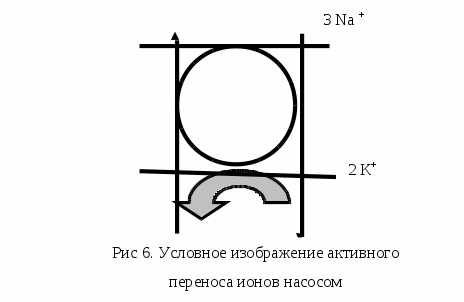
Натрий-калиевый насос работает, потребляя энергию АТФ, основным его компонентом является фермент – мембранная Nа, К – АТФаза. Движение ионов, по сути, главный механизм формирования МПП. Но в образовании МПП участвует добавочный механизм – прямой электрогенный эффект натрий-калиевого насоса (Рис.6). Этот эффект зависит от поляризации мембраны, возникающей за счет неравенства числа ионов Nа+ и К+. Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя. В самой мембране МПП проявляется как электрическое поле значительной напряженности, которое обеспечивает закрытое и открытое состояние ворот натриевые каналов, обеспечивающих состояние покоя и готовности к возбуждению.
Потенциал действия или потенциал возбуждения нервных клеток возникает в ответ на достаточное по силе раздражение. Это очень быстрый, кратковременный электрический процесс, который проявляется, в первую очередь, изменением электрического состояния клеточной мембраны. Раздражение способствует перезарядке, реверсии, т.е. заряд мембраны уменьшается от нуля и приобретает отрицательное значение: внутренняя сторона мембраны заряжается положительно, наружная – отрицательно. Затем вновь восстанавливается первоначальный заряд. Колебания потенциала, возникающее в результате кратковременной перезарядке мембраны и последующего восстановления ее исходного заряда, называется потенциалом действия. В потенциале действия различают пик (длительность 0,5 – 0,8 мс) и следовые потенциалы.
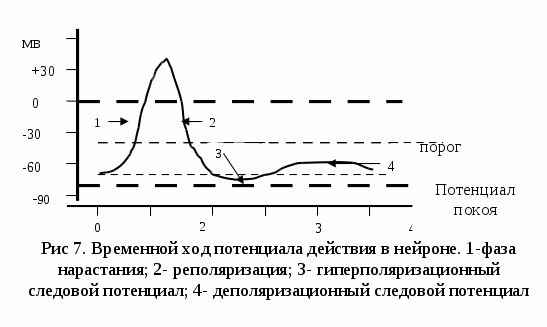
Современная мембранная теория объясняет механизм возникновения потенциала действия прохождением потоков ионов калия и натрия через каналы. Когда деполяризация достигает критической величины, за доли секунды, одновременно открываются максимальное количество натриевых каналов, проницаемость мембраны для ионов натрия увеличивается по сравнению с покоем в 500 раз. Ионы натрия устремляются внутрь клетки, унося с наружной поверхности мембраны положительный заряд, благодаря чему положительно заряженной становится внутренняя поверхность мембраны. Наружная же поверхность мембраны становится отрицательно заряженной. Но такое положение длится всего 0,5 – 1,5 мс. Количество открытых натриевых каналов уменьшается, проницаемость для ионов натрия уменьшается до исходного уровня, этот процесс называется инактивацией механизма натриевой проницаемости. Но во время инактивации механизма натриевой проницаемости увеличивается проницаемость мембраны для ионов калия. Выходя из клетки, ионы калия выносят положительный заряд, чем восстанавливают исходную разность потенциалов и мембрана опять приобретает снаружи положительный заряд, изнутри – отрицательный.
Потенциал действия, возникнув в одном участке нервной клетки, нервного или мышечного волокна, быстро распространяется по всей ее поверхности. Потенциал действия по силе превышает в 5-10 раз силу раздражения, способную вызвать возбуждение, это необходимо для возбуждения участка клетки, находящегося в покое. Между возбужденным участком и находящимися в состоянии покоя, возникает разность потенциалов, приводящая к возникновению круговых токов.
Скорость проведения возбуждения в волокнах скелетных мышц достигает 12-15 м/с, гладких мышц – 2-15, в безмякотных нервных волокнах – 0,5 – 3, в мякотных – 70-120 м/с.
Дата добавления: 2015-09-28; просмотров: 2739;