Доказательства генетического определения признаков пола
Важным доказательством в пользу наследственной детерминированности половой принадлежности организмов является наблюдаемое у большинства видов соотношение по полу 1:1 (табл. 6.1).
Таблица 6.1.Соотношение по полу у разных видов
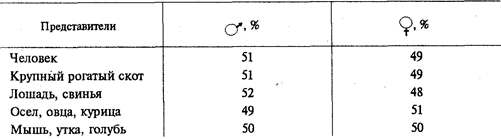
Такое соотношение может быть обусловлено образованием двух видов гамет представителями одного пола (гетерогаметный пол) и одного вида гамет — особями другого пола (гомогаметный пол). Это соответствует различиям в кариотипах организмов разных полов одного и того же вида, проявляющимся в половых хромосомах. У гомогаметного пола, имеющего одинаковые половые хромосомыXX,все гаметы несут гаплоидный набор аутосом плюс Х-хромосому. У гетерогаметного пола в кариотипе кроме аутосом содержатся две разные или только одна половая хромосома (XY или ХО). Его представители образуют два вида гамет, различающиеся по гетерохромосомам: Х и Y или Х и 0.
У разных видов организмов хромосомный механизм определения пола реализуется по-разному. У человека и других млекопитающих, а также у дрозофилы гомогаметным является женский пол (XX), а гетерогаметным — мужской (XY). У некоторых насекомых (клопы рода Protenor) гетерогаметный мужской пол имеет лишь одну Х-хромосому (ХО). У птиц и некоторых насекомых женский пол является гетерогаметным (XY), а мужской — гомогаметным (XX). У некоторых бабочек гетерогаметный женский пол имеет одну Х-хромосому (ХО). В большинстве выше описанных случаев пол вновь образующегося организма определяется сочетанием половых хромосом, возникающим в зиготе при оплодотворении (табл. 6.2).
Таблица 6.2.Хромосомный механизм определения полов
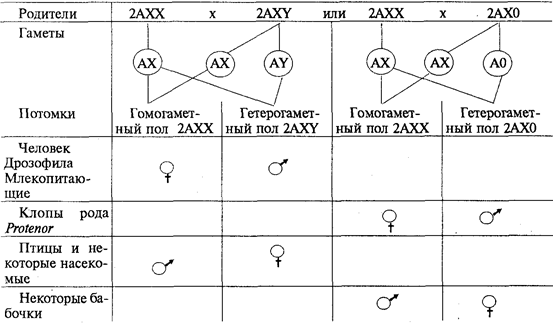
Таким образом, хромосомный механизм определения половой принадлежности организмов обеспечивает равновероятность встречаемости представителей обоих полов. Это имеет большой биологический смысл, так как обусловливает максимальную вероятность встречи самки и самца, потомки получают более разнообразную наследственную информацию, поддерживается оптимальная численность особей в популяции.
Исследования, проведенные на дрозофиле, обнаружили незначительную роль Y-хромосомы в определении пола. Особи с кариотипом ХО внешне являются типичными самцами, но стерильными, а особи с кариотипом XXY — плодовитые самки. Y-хромосома у дрозофилы по размеру близка к Х-хромосоме, однако она генетически инертна, так как состоит в основном из гетерохроматина. В связи с этим первичное определение пола у дрозофилы обусловлено не столько сочетанием в зиготе Х- и Y-хромосом, сколько соотношением числа Х-хромосом и наборов аутосом (половой индекс). У нормальных самок половой индекс равен 1 (2Х:2А), у нормальных самцов — 0,5 (XY:2A). Увеличение полового индекса более 1 при увеличении дозы Х-хромосомы (ЗХ:2А) приводит к развитию сверхсамок, а величина этого показателя ниже 0,5 (XY:3A) обеспечивает появление самцов. При значениях полового индекса более 0,5, но менее 1 (2Х:ЗА) развиваются интерсексы.
О значении дозы Х-хромосом в определении пола у дрозофилы свидетельствует развитие латерального гинандроморфизма у мух при утрате Х-хромосомы одной из клеток на стадии первого деления зиготы. В результате половина тела, развивающаяся из клетки с нормальным кариотипом (2АХХ), несет признаки самки, а другая половина, клетки которой лишены одной Х-хромосомы (2АХО), имеет признаки самца (рис. 6.2).
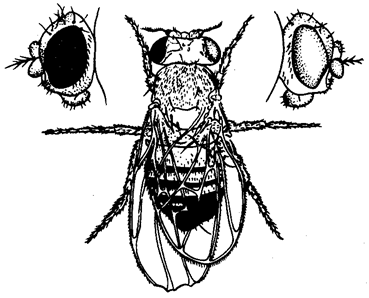
Рис. 6.2. Латеральный гинадроморфизм у Drosophila melanogaster, гетерозиготной по гену окраски глаз, находящемуся в Х-хромосоме
Левая половина мухи, клетки которой несут 2Х, имеет признаки самки и красный глаз; правая половина мухи, клетки которой несут одну Х-хромосому, имеет признаки самца и белый глаз
Обнаружение зависимости половой принадлежности развивающегося организма от дозы Х-хромосом у дрозофилы и некоторых других насекомых привело американского генетика С. Бриджеса (1922) к формулировке гипотезы генного баланса, в соответствии с которой организм изначально бисексуален, т.е. несет в себе задатки и мужского, и женского полов. Развитие признаков одного из них в ходе онтогенеза определяется балансом женских и мужских генов — детерминаторов пола. У дрозофилы эти гены сосредоточены не только в половой Х-хромосоме, но и в аутосомах. Поэтому пол организма у них зависит от соотношения этих хромосом. У плодовой мухи Y-хромосома, будучи генетически инертной, в определении признаков пола не участвует.
У человека Y-хромосома играет важную роль в детерминации пола. Она содержит определенное количество генов, часть из которых гомологична генам Х-хромосомы, а часть не имеет в ней гомологов и наследуется только по мужской линии. Некоторые из этих генов непосредственно связаны с детерминацией мужского пола. Поэтому у человека присутствие Y-хромосомы в кариотипе независимо от количества Х-хромосом (2AXXY, 2AXXXY) обеспечивает развитие мужского пола. Особи с кариотипом 2АХО являются женщинами, несмотря на уменьшение дозы Х-хромосомы. Однако значение баланса генов в определении половой принадлежности организма подтверждается тем, что особи с кариотипами 2АХО, 2АХХХ, 2AXXY, 2AXXXY и т.д. отличаются наличием пороков развития и часто стерильны.
Как было отмечено, у большинства видов животных пол организма определяется сочетанием или количеством половых хромосом в диплоидном наборе аутосом. Однако у некоторых насекомых (пчелы, муравьи, осы) представители разных полов отличаются числом наборов хромосом. Самки у них развиваются из оплодотворенных яиц и поэтому диплоидны, самцы — из неоплодотворенных яиц и первично гаплоидны. В такой ситуации соотношение по полу зависит от того, происходит оплодотворение или нет. Половой состав популяции может быть различным в разные периоды ее жизни.
Зависимость половой принадлежности организмов от сочетания или количества половых хромосом в его кариотипе свидетельствует о том, что гены, определяющие пол, располагаются главным образом в этих хромосомах. Однако известен целый ряд примеров, указывающих на то, что в развитии признаков пола принимают участие также гены, локализованные в аутосомах. В частности, в геноме дрозофилы присутствует ген, называемый трансформатором. Рецессивный аллель этого гена в гомозиготном состоянии tt превращает женские зиготы 2Х в фенотипических самцов, которые оказываются стерильными. Самки1 XXt+t+ или XXt+t плодовиты, особи с кариотипом XYtt — нормальные самцы, организмы с кариотипом XXtt — превращенные стерильные самцы. Другим примером влияния аутосомных генов на развитие признаков пола может служить существование у кукурузы генов, мутантные аллели которых превращают ее из однодомного растения в двудомное, препятствуя развитию органов либо мужского, либо женского пола. Рецессивный аллель гена sk(silkless) в гомозиготном состоянии вызывает недоразвитие семяпочек и превращает растение в мужское. Мутантный аллель другого гена ts (tassel seed) в гомозиготном состоянии вызывает развитие семяпочек и семян в метелке, а пыльники не развиваются.
У человека развитие организма по мужскому типу обеспечивается не только геном, расположенным в Y-хромосоме и определяющим способность к синтезу мужского полового гормона — тестостерона, но и Х-сцепленным геном, контролирующим синтез белка-рецептора этого гормона. Мутация упомянутого Х-сцепленного гена приводит к развитию синдрома тестикулярной феминизации (см. разд. 3.6.5.2).
Приведенные примеры доказывают, что пол организмов как признак развивается на основе полученной от родителей наследственной информации и находится под контролем группы взаимодействующих генов, расположенных как в половых хромосомах, так и в аутосомах.
Дата добавления: 2015-09-07; просмотров: 1415;