Цитоплазма (cytoplasma).
Структурними компонентами цитоплазми є гіалоплазма, органели і включення.
Гіалоплазма— найрідша частина цитоплазми, в якій містяться органели і включення. У загальному об'ємі цитоплазми гіалоплазма становить близько 50 %. Вона включає цитозоль (воду з розчиненими у ній неорганічними та органічними речовинами) і цитоматрикс (трабекулярну сітку волокон білкової природи товщиною 2... З нм).
Органели— постійні структури цитоплазми, які мають певну будову і виконують спеціалізовану функцію. Органели поділяються на мікроскопічні, видимі під світловим мікроскопом, і субмікроскопічні, які можна побачити лише з допомогою електронного мікроскопа. За наявністю у складі органел біологічної мембрани їх поділяють на мембранні та немембранні. До мембранних органел належать: мітохондрії, лізосоми, пероксисоми, ендоплазматична сітка, комплекс Гольджі. Немембранними органелами є рибосоми, мікрофіламенти, мікротрубочки, центросома (клітинний центр). Ці дев'ять органел називають органелами загального призначення, оскільки вони присутні у всіх без виключення видах клітин. Органели загального призначення можуть утворювати характерні конгломерати у цитоплазмі клітин. Такі конгломерати з переважним розвитком і особливою організацією органел того чи іншого виду називають спеціальними органелами (тонофібрили клітин епітелію, міофібрили м'язових клітин і волокон, нейрофібрили нервових клітин та деякі інші).
Мітохондрії — мікроскопічні мембранні органели загального призначення, основна функція яких — утворення необхідної для життєдіяльності клітини енергії та нагромадження її у складі молекул аденозинтрифосфорної кислоти (АТФ). Крім цього, мітохондрії беруть участь у регуляції обміну води, депонуванні іонів кальцію, продукції попередників стероїдних гормонів. Мітохондрії відкриті німецьким дослідником Ф. Альтманом у кінці XIX століття. Під світловим мікроскопом мітохондрії мають вигляд дрібних крапочок і ниточок завтовшки близько 0,5 мкм і довжиною 1... 10 мкм. За допомогою електронного мікроскопа у складі кожної мітохондрії, яка має неправильну овальну або витягнуту форму, можна розрізнити дві мембрани: зовнішню гладку і внутрішню складчасту, що утворюють вирости (кристи) всередину мітохондрії (рис. 1.4 і Див. рис. 1.1). Всередині мітохондрія заповнена електронно-щільною речовиною, яка називається матриксом. У матриксі, а також у внутрішній мембрані мітохондрій містяться білки-ферменти, які забезпечують синтез АТФ шляхом окисного фосфорилювання аденозиндифосфату (АДФ). Мітохондрії – єдині органели клітини, в яких знайдені молекули власної дезоксирибонуклеїнової кислоти (ДНК);до їх матриксу входять також різні види РНК та рибосоми.
Лізосоми — субмікроскопічні мембранні органели загального призначення, відкриті у 1955 р. Христіаном де Дювом. Основна функція лізосом — розщеплення біополімерів різного хімічного складу (так зване клітинне травлення). Для цього у лізосомах міститься набір гідролітичних ферментів (зараз їх відомо понад 60). Маркерним (визначальним) ферментом лізосом є кисла фосфатаза. Ферментні комплекси матриксу лізосоми знаходяться у замкненому мембранному мішечку діаметром близько 0,2...0,4 мкм, який перешкоджає попаданню лізосомних ферментів у гіалоплазму і запобігає самоперетравленню клітини (рис. 1.5 і див. рис. 1.1). Залежно від ультраструктурних і функціональних особливостей лізосом їх поділяють на первинні (ферменти яких знаходяться у неактивному стані), вторинні, або фагосоми (активовані ферменти в них безпосередньо контактують з розщеплюваними біополімерами), а також залишкові тільця (оточені біомембраною нерозщеплені залишки). Слід зауважити, що лізосоми можуть брати участь як у розщепленні власних макромолекулярних комплексів клітини (таке явище носить назву аутофагоцитозу), так і в перетравлюванні поглинутих клітиною частинок (гетерофагоцитоз). Недостатність того чи іншого лізосомного ферменту призводить до нагромадження в клітині аномальних біополімерів, що зумовлює розвиток так званих лізосомних хвороб нагромадження (тезаурісмозів). До цього часу описано понад 30 різних лізосомних хвороб нагромадження.
 Пероксисоми— субмікроскопічні мембранні органели загального призначення, відкриті на початку 60-х років спільними зусиллями біохіміків і морфологів. Пероксисомам належить вирішальна роль у процесах детоксикації клітини (позбавленні її від дії токсичних продуктів обміну речовин). Утворений біомембраною мішечок округлої форми діаметром 0,2...0,5 мкм заповнений ферментами (матриксом), серед яких маркерним є каталаза. У центрі матриксу пероксисом ) за допомогою електронного мікроскопа знайдена щільна серцевина (кристалоїд), яка містить волокнисті та трубчасті макро-молекулярні утвори. Ферментні системи пероксисом спрямовані на утилізацію хімічно активного атомарного кисню (у першу чергу шляхом регуляції обміну і розщеплення перекису водню), а також забезпечують розщеплення етилового спирту, сечової кислоти, регуляцію обміну ліпідів.
Пероксисоми— субмікроскопічні мембранні органели загального призначення, відкриті на початку 60-х років спільними зусиллями біохіміків і морфологів. Пероксисомам належить вирішальна роль у процесах детоксикації клітини (позбавленні її від дії токсичних продуктів обміну речовин). Утворений біомембраною мішечок округлої форми діаметром 0,2...0,5 мкм заповнений ферментами (матриксом), серед яких маркерним є каталаза. У центрі матриксу пероксисом ) за допомогою електронного мікроскопа знайдена щільна серцевина (кристалоїд), яка містить волокнисті та трубчасті макро-молекулярні утвори. Ферментні системи пероксисом спрямовані на утилізацію хімічно активного атомарного кисню (у першу чергу шляхом регуляції обміну і розщеплення перекису водню), а також забезпечують розщеплення етилового спирту, сечової кислоти, регуляцію обміну ліпідів.
 Ендоплазматична сітка –субмікроскопічна мембранна органела загального призначення, яка утворює єдину внутріш-ньоцитоплазматичну циркуляційну систему, вперше описану К. Портером у 1945 р. Вона є замкненою сукупністю канальців, мішечків та цистерн, утворених суцільною (неперервною) біомембраною (див. рис. 1.1, 1.4 і 1.8). Мембрана ендоплазматичної сітки безпосередньо контактує з плазмолемою клітини і з мембранами ядра. Розрізняють гладку і гранулярну (шорстку) ендоплазматичні сітки. Гладка ендоплазматична сітка, діаметр канальців якої 50... 100 нм, утворена лише подвоєною мембраною. Гранулярна ендоплазматична сітка утворена подвоєною біомембраною, до якої з боку гіалоплазми прикріплені рибосоми (рис. 1.8). Діаметр канальців гранулярної сітки від 20 до 1000 нм. Функція гладкої ендоплазматичної сітки пов'язана з метаболізмом ліпідів і вуглеводів,детоксикацією шкідливих для клітини хімічних сполук, а також депонуванням іонів кальцію. Функція гранулярної ендоплазматичної сітки зумовлена присутністю рибосом і полягає у біосинтезі білків як для потреб самої клітини, так і для виведення у зовнішнє середовище. Крім виконання метаболічної та циркуляторної ролей, ендоплазматична сітка — єдина органела, в якій відбувається новоутвір мембранних структур клітини. Синтезовані гранулярною ендоплазматичною сіткою, компоненти біомембрани можуть включатися до складу мішечків лізосом, пероксисом, елементів комплексу Гольджі, плазмолеми, оболонки ядра, а також використовуватися для самовідтворення елементів ендоплазматичної сітки.
Ендоплазматична сітка –субмікроскопічна мембранна органела загального призначення, яка утворює єдину внутріш-ньоцитоплазматичну циркуляційну систему, вперше описану К. Портером у 1945 р. Вона є замкненою сукупністю канальців, мішечків та цистерн, утворених суцільною (неперервною) біомембраною (див. рис. 1.1, 1.4 і 1.8). Мембрана ендоплазматичної сітки безпосередньо контактує з плазмолемою клітини і з мембранами ядра. Розрізняють гладку і гранулярну (шорстку) ендоплазматичні сітки. Гладка ендоплазматична сітка, діаметр канальців якої 50... 100 нм, утворена лише подвоєною мембраною. Гранулярна ендоплазматична сітка утворена подвоєною біомембраною, до якої з боку гіалоплазми прикріплені рибосоми (рис. 1.8). Діаметр канальців гранулярної сітки від 20 до 1000 нм. Функція гладкої ендоплазматичної сітки пов'язана з метаболізмом ліпідів і вуглеводів,детоксикацією шкідливих для клітини хімічних сполук, а також депонуванням іонів кальцію. Функція гранулярної ендоплазматичної сітки зумовлена присутністю рибосом і полягає у біосинтезі білків як для потреб самої клітини, так і для виведення у зовнішнє середовище. Крім виконання метаболічної та циркуляторної ролей, ендоплазматична сітка — єдина органела, в якій відбувається новоутвір мембранних структур клітини. Синтезовані гранулярною ендоплазматичною сіткою, компоненти біомембрани можуть включатися до складу мішечків лізосом, пероксисом, елементів комплексу Гольджі, плазмолеми, оболонки ядра, а також використовуватися для самовідтворення елементів ендоплазматичної сітки.
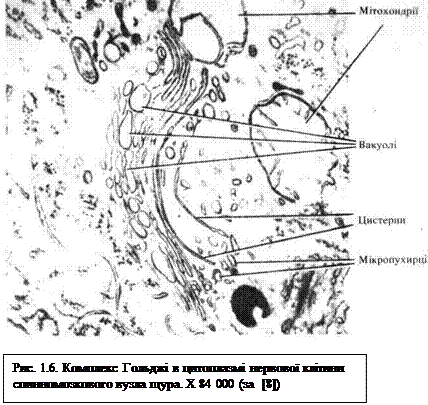
 Комплекс Гольджі— мікроскопічна мембранна органела загального призначення, у якій завершується процес формування продуктів синтетичної діяльності клітини, зокрема здійснюється їх кінцеве глікозування. Комплекс Гольджі нагромаджує секреторні речовини і забезпечує їх виведення за межі клітини. Органела названа іменем італійського гістолога Камілло Гольджі, який у 1898 р. описав цей комплекс у складі нервових клітин. Морфологічно комплекс Гольджі — це сукупність пов'язаних між собою цистерн товщиною близько 25 нм, сплющених у центральній частині і розширених на периферії. Проміжки між окремими цистернами становлять 20...25 нм. Від розширених країв цистерн відшнуровуються дрібні пухирці (рис; 1.6 і див. рис. 1.1). Окрема сукупність таких цистерн і пухирців носить назву диктіосоми. Образно диктіосому порівнюють зі стосом тарілок, повернутих ввігнутим боком до ядра. В одній клітині може бути кілька диктіосом, відокремлених одна від одної прошарками гіалоплазми. Завершується синтез (дозрівання) секреторних продуктів при переміщенні їх з цистерн, розташованих поблизу ядра (так звана цис-локалізація цистерн), у напрямку плазмолеми (транс-локалізація цистерн). Пухирці, які відшнуровуються від країв цистерн, містять сформовані, готові до виведення з клітини секреторні продукти. Нагромаджені у складі пухирців макромолекулярні комплекси виводяться шляхом вмонтовування мембран пухирців у плазмолему, або виштовхуванням сформованих (зрілих) пухирців за межі клітини.
Комплекс Гольджі— мікроскопічна мембранна органела загального призначення, у якій завершується процес формування продуктів синтетичної діяльності клітини, зокрема здійснюється їх кінцеве глікозування. Комплекс Гольджі нагромаджує секреторні речовини і забезпечує їх виведення за межі клітини. Органела названа іменем італійського гістолога Камілло Гольджі, який у 1898 р. описав цей комплекс у складі нервових клітин. Морфологічно комплекс Гольджі — це сукупність пов'язаних між собою цистерн товщиною близько 25 нм, сплющених у центральній частині і розширених на периферії. Проміжки між окремими цистернами становлять 20...25 нм. Від розширених країв цистерн відшнуровуються дрібні пухирці (рис; 1.6 і див. рис. 1.1). Окрема сукупність таких цистерн і пухирців носить назву диктіосоми. Образно диктіосому порівнюють зі стосом тарілок, повернутих ввігнутим боком до ядра. В одній клітині може бути кілька диктіосом, відокремлених одна від одної прошарками гіалоплазми. Завершується синтез (дозрівання) секреторних продуктів при переміщенні їх з цистерн, розташованих поблизу ядра (так звана цис-локалізація цистерн), у напрямку плазмолеми (транс-локалізація цистерн). Пухирці, які відшнуровуються від країв цистерн, містять сформовані, готові до виведення з клітини секреторні продукти. Нагромаджені у складі пухирців макромолекулярні комплекси виводяться шляхом вмонтовування мембран пухирців у плазмолему, або виштовхуванням сформованих (зрілих) пухирців за межі клітини.
Рибосоми— субмікроскопічні немембранні органели загального призначення, у яких амінокислоти сполучаються, утворюючи пептидний ланцюг, тобто синтезуються білкові молекули. Вперше рибосоми у складі гранулярної ендоплазматичної сітки описав Дж. Паладе. Морфологічно у рибосомах налічуються дві субодиниці. їх сполучення утворює структуру, яка за формою нагадує гриб (див. рис. 1.1). Діаметр рибосом — близько 20 нм. З хімічної точки зору рибосома — це рибонуклеопротеїновий комплекс рибосомної РНКі білка у співвідношенні 1:1. При дії пошкоджуючих факторів або порушенні електролітного гомеостазу клітини (брак іонів магнію) спостерігається розпад рибосоми на субодиниці (дезагрегація), біологічна активність її при цьому втрачається. Кілька рибосом, «нанизаних» на спільну нитку інформаційної РНК,називаються полісомами. На завислих у гіалоплазмі полісомах здійснюється синтез білків переважно для внутрішніх потреб клітини. Полісоми, пов'язані з мембранами ендоплазматичної сітки, переважно синтезують білки для виведення за межі клітини.
 Мікрофіламенти— субмікроскопічні немембранні органели загального призначення, які виконують роль цитоскелету, а також скоротливого апарату клітини. Власне мікрофіламенти — це тонкі волоконця діаметром близько 5 нм, побудовані з білків актину, міозину, тропо-міозину або альфа-актиніну. Вони розміщені переважно в кортикальній (підмембранній) зоні клітини та у складі її цитоплазматичних виростів. їхня функція — скоротливо-рухова.
Мікрофіламенти— субмікроскопічні немембранні органели загального призначення, які виконують роль цитоскелету, а також скоротливого апарату клітини. Власне мікрофіламенти — це тонкі волоконця діаметром близько 5 нм, побудовані з білків актину, міозину, тропо-міозину або альфа-актиніну. Вони розміщені переважно в кортикальній (підмембранній) зоні клітини та у складі її цитоплазматичних виростів. їхня функція — скоротливо-рухова.
Друга група мікрофіламентів (так звані проміжні) має діаметр 10... 15 нм. Білок, з якого побудовані проміжні мікрофі-ламенти, є суто специфічною ознакою клітин того чи іншого типу. Наприклад, кератин є гістохімічним маркером клітин епітелію, віментин — сполучної тканини, д є с м і н — м'язів, білок нейрофіламентів і гліальний фібрилярний білок — нервової тканини. Проміжні мікрофіламенти в основному відповідають за збереження клітиною своєї форми. В останні роки підтверджений їх зв'язок з регуляцією активності геному і процесами клітинної диференціації. Слід зауважити, що у складі спеціалізованих клітин мікрофіламенти можуть утворювати своєрідні пучки складнішої будови (тонофібрили епітеліальних клітин, міофібрили м'язів, нейрофібрили нервових клітин). Внаслідок особливої організації тонофібрили, міофібрили та нейрофібрили належать до спеціальних оргапел відповідних видів клітин.
Мікротрубочки— субмікроскопічні немембранні органели загального призначення, основна функція яких полягає у забезпеченні рухливості клітинних органел, а також в утворенні цитоскелету (див. рис. 1.1). Мікротрубочки побудовані з глобулярних білків тубулінів, молекули яких здатні до полімеризації. Особливим способом «нанизуючись» одна на одну, окремі молекули утворюють своєрідні «бусинки»; 13 ниток бусинок, які розміщені паралельно, формують порожнистий циліндр діаметром близько 25 нм з внутрішнім просвітом 15 нм. Товщина стінки циліндра відповідає діаметру однієї молекули тубуліну і становить 5 нм. Полімеризація молекул тубулінів є динамічним процесом, який припиняється під дією несприятливих факторів зовнішнього середовища (пониження температури, обробка колхіцином). Часткова деполімеризація мікротрубочок призводить до їх вкорочення, повна — до розпаду (дисоціації) на окремі молекули тубулінів. Мікротрубочки є основою будови центросоми, а також таких спеціалізованих структур як війки та джгутики.
Центросома— (клітинний центр) — мікроскопічна немембранна органела загального призначення, описана В. Флемінгом у 1875 р., яка забезпечує розходження хромосом при поділі клітини. Ця органела знайдена в усіх клітинах тварин і людини за виключенням яйцеклітин. У клітині, що не готується до. поділу, центросома розміщена біля ядра і складається з двох повністю сформованих ц є н т р і о-л є й, оточених центросферою. Дві розташовані поряд центріолі називають диплосомою. Кожна центріоля у своїй основі містить дев'ять триплетів паралельно орієнтованих мікротрубочок, які у просторовому зображенні формують циліндр діаметром 200 нм і довжиною близько 500 нм (див. рис. 1.1). Крім мікротрубочок, до складу центріолі входять специфічні макромолекулярні утвори, так звані «ручки», за допомогою яких триплети пов'язані між собою. У складі «ручок» міститься білок динеїн, який має АТФ-азну активність і якому належить вирішальна роль у механізмах реалізації рухових функцій центріолей. Довгі осі обох центріолей розташовані у взаємно перпендикулярних площинах. Центросфера являє собою позбавлену органел гіалоплазму навколо центріолей, яку у радіальному напрямі пронизують мікрофіламенти і мікротрубочки. При підготовці клітини до поділу відбувається подвоєння (дуплікація) центріолей з наступним розход- женням кожної новоутвореної пари до полюсів клітини. Основою функціональної активності центросоми є механізм стимулювання полімеризації тубулінів, що зумовлює ріст існуючих і утворення нових мікротрубочок.
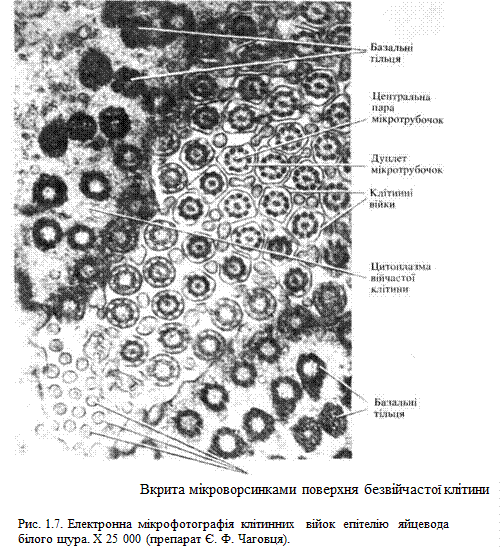 Війки(рис. 1.7) і джгутики— це тонкі вирости цитоплазми, основою яких є високоорганізована система мікротрубочок. Довжина війок становить 5…10 мкм, джгутиків — досягає 150 мкм. Діаметр обох цих структур приблизно однаковий і становить близько 200 нм. Зовні війку покриває плазмолема, всередині її проходить осьова нитка — аксонема. Аксонема містить дев'ять пар (дуплетів) мікротрубочок, які формують порожнистий циліндр діаметром близько 150 нм. У центрі цього циліндра локалізується центральна пара мікротрубочок. Систему мікротрубочок аксонеми описують формулою (9X2)+2. Біля основи війки в ділянці її переходу в цитоплазму лежить так зване базальне тільце, яке за будовою нагадує центріолю і складається з дев'яти триплетів мікротрубочок. Формула системи мікротрубочок базального тільця має вигляд (9X3)+0. Часто біля основи війки лежать два базальні тільця, довгі осі яких розміщені під прямим кутом одна до одної. Базальне тільце і аксонема пов'язані між собою: дві мікротрубочки з кожного триплету базального тільця продовжуються у дуплет мікротрубочок аксонеми. Незначні зміщення дуплетів мікротрубочок можуть викликати вигинання всієї війки або джгутика. У різних клітинах рух війок і джгутиків може нагадувати рух маятника, бути лійкоподібним, хвилеподібним тощо. Вільні клітини, що мають війки і джгутики, можуть переміщуватися у просторі (наприклад, сперматозоїди). Фіксовані клітини з війками на апікальній поверхні рухом своїх війок можуть транспортувати рідину або окремі частинки (наприклад, війчасті клітини дихальних шляхів, маткових труб тощо).
Війки(рис. 1.7) і джгутики— це тонкі вирости цитоплазми, основою яких є високоорганізована система мікротрубочок. Довжина війок становить 5…10 мкм, джгутиків — досягає 150 мкм. Діаметр обох цих структур приблизно однаковий і становить близько 200 нм. Зовні війку покриває плазмолема, всередині її проходить осьова нитка — аксонема. Аксонема містить дев'ять пар (дуплетів) мікротрубочок, які формують порожнистий циліндр діаметром близько 150 нм. У центрі цього циліндра локалізується центральна пара мікротрубочок. Систему мікротрубочок аксонеми описують формулою (9X2)+2. Біля основи війки в ділянці її переходу в цитоплазму лежить так зване базальне тільце, яке за будовою нагадує центріолю і складається з дев'яти триплетів мікротрубочок. Формула системи мікротрубочок базального тільця має вигляд (9X3)+0. Часто біля основи війки лежать два базальні тільця, довгі осі яких розміщені під прямим кутом одна до одної. Базальне тільце і аксонема пов'язані між собою: дві мікротрубочки з кожного триплету базального тільця продовжуються у дуплет мікротрубочок аксонеми. Незначні зміщення дуплетів мікротрубочок можуть викликати вигинання всієї війки або джгутика. У різних клітинах рух війок і джгутиків може нагадувати рух маятника, бути лійкоподібним, хвилеподібним тощо. Вільні клітини, що мають війки і джгутики, можуть переміщуватися у просторі (наприклад, сперматозоїди). Фіксовані клітини з війками на апікальній поверхні рухом своїх війок можуть транспортувати рідину або окремі частинки (наприклад, війчасті клітини дихальних шляхів, маткових труб тощо).
Включення на відміну від органел не є постійними структурними компонентами цитоплазми і не мають строго визначеної будови. Включення бувають екзо- та ендогенні. Останні залежно від їхнього функціонального призначення поділяють на екскреторні, трофічні, пігментні тощо. Власне кажучи, включення — це макромолекулярні конгломерати, які клітина у тих чи інших умовах свого функціонування нагромаджує у цитоплазмі.
Дата добавления: 2015-09-18; просмотров: 2124;