ЯДРО. РЕПРОДУКЦІЯ КЛІТИН
Ядро (nucleus) — важлива складова частина клітини. Разом з цитоплазмою утворює єдину інтегровану систему, яка знаходиться у стані динамічної рівноваги. Клітина не може довго існувати без ядра (швидко гине при його видаленні — енуклеації), але і ядро теж без цитоплазми не здатне до самостійного життя. Термін «ядро» належить Р. Броуну, який вперше застосував його у 1833 р. при описанні рослинних клітин.
Ядро виконує дві групи загальних функцій. Перша пов'язана зі збереженням генетичної (спадкової) інформації серед клітинних поколінь. Це такі функції: підтримання постійної структури ДНК за допомогою так званих репараційних ферментів, які можуть відновити молекулу ДНК після її ушкоджень (у тому числі, радіаційних); редуплікація молекул ДНК (тобто якісне і кількісне подвоєння генетичного матеріалу); розподіл генетичного матеріалу між дочірніми клітинами під час мітозу; рекомбінація генетичного матеріалу в процесі мейозу. Друга група ядерних функцій стосується реалізації генетичної інформації, тобто полягає у створенні апарату білкового синтезу. До цих функцій належать синтез усіх видів РНК (інформаційної, транспортної, рибосомної), а також побудова рибосом.
Усі клітини людського організму містять ядро, за винятком високоспеціалізованих клітин крові — еритроцитів, що втрачають ядро у процесі свого розвитку і є без'ядерними. Переважна більшість клітин містить одне ядро, але бувають двоядерні клітини (20 % клітин печінки є двоядерними), а також багатоядерні (наприклад,остеокласти — клітини кісткової тканини).
За формою ядра найчастіше бувають сферичні, але можуть мати й іншу форму — паличкоподібну, бобовидну, кільцеподібну, сегментовану. Форма ядра залежить від форми клітини (видовжені клітини гладких м'язів мають видовжене паличкоподібне ядро); від кількості включень (ядро жирової клітини набуває сплющеної форми під впливом великої жирової краплі, що займає майже всю клітину); розташування органел (форма ядра моноцита бобовидна завдяки розташуванню в місці його заглибини центросоми).
Ядро локалізується завжди у певному місці клітини. Наприклад, у циліндричних клітинах шлунка, кишки воно займає базальне положення. Розміри ядер від 3—4 до 40 мкм. Кожний тип клітини має своє постійне співвідношення між об'ємом ядра і цитоплазми. Ця константа носить назву індексуГертвіг а. Згідно значень цього індексу клітини поділяють на ядерні (з великим індексом Гертвіга) та цитоплазматичні (з малим індексом Гертвіга).
Ядро може бути в двох станах — мітотичному (під час поділу) та інтерфазному (між поділами). Останнє називають також метаболічним ядром, що підкреслює його функціональний стан. У живій клітині інтерфазне ядро виглядає оптично пустим, видно лише ядерце. Структури ядра у вигляді ниточок, зерняток у живій клітині можна спостерігати лише при дії на неї пошкоджуючих агентів, коли клітина переходить у стан так званого паранекрозу (стан на межі життя і смерті). З цього стану клітина може або повернутися до нормальної життєдіяльності, або загинути. Морфологічно розрізняють такі зміни ядра при загибелі клітини: каріопікноз (ущільнення), каріорексис (розпад), каріолізис (розчинення).
На фіксованому і забарвленому препараті у складі інтерфаз-ного ядра розрізняють ядерну оболонку, хроматин, ядерце та каріоплазму (рис. 1.8, 1.9).
Ядерна оболонка.Ядра всіх еукаріотичних клітин оточені оболонкою (нуклеолемою). Ядерна оболонка складається з двох біологічних мембран — зовнішньої і внутрішньої, відокремлених перинуклеарним простором шириною 20...60 нм. Кожна з мембран має товщину 7...8 нм і морфологічно подібна до інших клітинних мембран. Таким чином, ядерна оболонка нагадує порожнистий двошаровий мішок, що відокремлює вміст ядра від цитоплазми. Зовнішня мембрана ядерної оболонки має ряд структурних особливостей, що дає змогу віднести її до мембранної системи ендоплазматичної сітки. Наприклад, на зовнішній ядерній мембрані здебільшого розташовується невелика кількість рибосом. Існує багато спостережень про безпосередній перехід зовнішньої ядерної мембрани у систему каналів гранулярної ендоплазматичної сітки. Зовнішня ядерна мембрана більшості тваринних і рослинних клітин не є рівною і може утворювати різного розміру вирости у вигляді пухирців або довгих трубчастих утворів у бік цитоплазми. Внутрішня ядерна мембрана пов'язана з хромосомним матеріалом ядра. 3 боку каріоплазми до внутрішньої ядерної мембрани прилягає так званий фібрилярний шар, побудований з фібрил; його, однак, знайдено не у всіх клітинах.
 Ядерна оболонка не є суцільною, її характерними структурами є специфічні отвори, що мають назву ядерних пор. Пори утворюються за рахунок злиття двох ядерних мембран і є округлими наскрізними перфораціями діаметром 80...90 нм (рис. 1.10). їх заповнюють складноорганізовані глобулярні та фібрилярні структури, які разом з мембранною перфорацією утворюють так званий комплекс пори. Останній побудований з трьох рядів гранул по вісім штук у кожному ряді; діаметр гранули 25 нм. Гранули розташовані на межі отвору в ядерній оболонці: один ряд лежить з боку ядра, другий — з боку цитоплазми, третій — у центральній частині пори. У центрі пори розташована центральна гранула. Від гранул відходять фібрилярні білкові структури, які сходяться в центрі, утворюючи перегородку поперек пори, так звану діафрагму пори. Розмір ядерних пор для кожного виду клітин є величиною сталою.
Ядерна оболонка не є суцільною, її характерними структурами є специфічні отвори, що мають назву ядерних пор. Пори утворюються за рахунок злиття двох ядерних мембран і є округлими наскрізними перфораціями діаметром 80...90 нм (рис. 1.10). їх заповнюють складноорганізовані глобулярні та фібрилярні структури, які разом з мембранною перфорацією утворюють так званий комплекс пори. Останній побудований з трьох рядів гранул по вісім штук у кожному ряді; діаметр гранули 25 нм. Гранули розташовані на межі отвору в ядерній оболонці: один ряд лежить з боку ядра, другий — з боку цитоплазми, третій — у центральній частині пори. У центрі пори розташована центральна гранула. Від гранул відходять фібрилярні білкові структури, які сходяться в центрі, утворюючи перегородку поперек пори, так звану діафрагму пори. Розмір ядерних пор для кожного виду клітин є величиною сталою.
Число ж пор на одиницю поверхні ядра може змінюватися залежно від функціонального стану клітини, її метаболічної активності: чим вона вища, тим більша густина пор на поверхні нуклеолеми.
Ядерна оболонка виконує ряд важливих функцій. Перша з них — бар'єрна функція: ядерна оболонка відокремлює вміст ядра, його генетичний матеріал від цитоплазми, обмежує вільний доступ в ядро та вихід із нього різних речовин. Друга функція — регуляція транспорту макромолекул між ядром і цитоплазмою. Наприклад, відомо, що гістони та інші негістонові білки після синтезу в цитоплазмі мігрують у ядро. Відомий також і зворотний процес транспорту речовин з ядра в цитоплазму. Це, у першу чергу, стосується транспорту РНК і РНП, що синтезуються виключно в ядрі. Механізм транспорту високомолекулярних сполук, а також рибосом через ядерну оболонку не зовсім зрозумілий, можливо, він здійснюється через пори.
Одна з суттєвих функцій ядерної оболонки — участь у створенні внутрішньоядерного порядку шляхом фіксації хромосомного матеріалу в інтерфазі до внутрішньої ядерної мембрани. Існують дані про переважний зв'язок з ядерною оболонкою гетерохроматинових ділянок інтерфазних хромосом. Ще з класичних цитологічних описів відомо про те, що частина хроматину локалізується на периферії ядра. Цей так званий периферійний хроматин структурно пов'язаний із внутрішньою ядерною мембраною. Крім периферійного хроматину, з ядерною оболонкою контактують прицентромерні, теломерні та ядерцеві ділянки гетерохроматину, статеві хромосоми тощо. Отже, можна вважати, що кожна деконденсована в інтерфазі хромосома «заякорена» на ядерній оболонці за допомогою гетерохроматинових ділянок і, таким чином, її положення стає фіксованим у просторі ядра.
 Хроматин. На фіксованому і забарвленому препараті в інтер-фазному ядрі видно зерна, грудочки, що добре забарвлюються основними барвниками. Цей компонент ядра вперше описав Вальтер Флемінг у 1881 р. і назвав хроматином (від грецького «хрома» — колір, барва). Хроматин — це основна структура інтерфазного ядра, яка зумовлює специфічний для кожного типу клітин хроматиновий малюнок ядра. Цей малюнок є ніби власною печаткою клітини, яка дає змогу пізнавати різні види клітин. Хроматин є структурним аналогом хромосом, які можна бачити лише під час мітозу. Хімічний склад хроматину такий, як і хромосом: основою є молекула ДНК, оточена білками-гістонами. Крім того, у хроматині виявлено невелику кількість РНК — продуктів процесу транскрипції. Співвідношення вказаних хімічних компонентів у хроматині ДНК:білок:РНК= = 1:1,3:0,2.
Хроматин. На фіксованому і забарвленому препараті в інтер-фазному ядрі видно зерна, грудочки, що добре забарвлюються основними барвниками. Цей компонент ядра вперше описав Вальтер Флемінг у 1881 р. і назвав хроматином (від грецького «хрома» — колір, барва). Хроматин — це основна структура інтерфазного ядра, яка зумовлює специфічний для кожного типу клітин хроматиновий малюнок ядра. Цей малюнок є ніби власною печаткою клітини, яка дає змогу пізнавати різні види клітин. Хроматин є структурним аналогом хромосом, які можна бачити лише під час мітозу. Хімічний склад хроматину такий, як і хромосом: основою є молекула ДНК, оточена білками-гістонами. Крім того, у хроматині виявлено невелику кількість РНК — продуктів процесу транскрипції. Співвідношення вказаних хімічних компонентів у хроматині ДНК:білок:РНК= = 1:1,3:0,2.
Морфологи розрізняють два види- хроматину: гетерохроматин і еухроматин. Перший відповідає конденсованим під час інтерфази ділянкам хромосом; він є функціонально неактивним. Цей хроматин добре забарвлюється, саме його можна бачити на гістологічному препараті. Гетерохроматин поділяється на структурний (це ділянки хромосом, що постійно конденсовані) та факультативний (може деконденсуватись і переходити в еухроматин). Еухроматин відповідає деконденсованим в інтерфазі ділянкам хромосом. Це робочий, функціонально активний хроматин. Він не забарвлюється, його не видно на гістологічному препараті. Під час мітозу весь еухроматин конденсується і входить до складу хромосом.
В окремих випадках ціла хромосома в період інтерфази може залишатися у конденсованому (тобто гетерохроматизованому) стані, мати вигляд грудочки гетерохроматину. Наприклад, одна з Х-хромосом у соматичних клітинах жіночого організму підлягає гетерохроматизації на початкових стадіях ембріогенезу (під час дроблення) і не функціонує. Вперше цей хроматин був описаний М. Барром і Л. Бертра-мом у 1949 р. і отримав назву статевого хроматину, або тілець Барра. У різних клітинах статевий хроматин має різний вигляд. Наприклад, у нейтрофільних лейкоцитах він виглядає як барабанна паличка, що виступає на поверхні одного із сегментів ядра. В епітеліальних клітинах слизової оболонки ротової порожнини статевий хроматин виглядає як добре помітна напівсферична грудочка гетерохроматину, прикріплена до внутрішньої ядерної мембрани. Той факт, що. статевий хроматин є не що  інше як гетерохроматизована одна з Х-хромосом, встановила вперше англійська дослідниця Мері Лайон, на честь якої процес переходу Х-хромосоми у стан гетерохроматину було названо лайонізацією. Визначення статевого хроматину використовується для встановлення генетичної статі організму (в судовій медицині, акушерстві), а також для встановлення кількості Х-хромосом у каріотипі індивіда (вона дорівнює кількості тілець статевого хроматину +1).
інше як гетерохроматизована одна з Х-хромосом, встановила вперше англійська дослідниця Мері Лайон, на честь якої процес переходу Х-хромосоми у стан гетерохроматину було названо лайонізацією. Визначення статевого хроматину використовується для встановлення генетичної статі організму (в судовій медицині, акушерстві), а також для встановлення кількості Х-хромосом у каріотипі індивіда (вона дорівнює кількості тілець статевого хроматину +1).
На ультраструктурному рівні в складі інтерфазного хроматину виявляються елементарні хромосомні фібрили товщиною 20...25 нм, які побудовані з фібрил товщиною 10 нм. Основу останніх становить молекула ДНК в комплексі з гістонами, що має вигляд намиста. Кожна намистина, що має назву нуклеосоми, складається з фрагменту подвійної спіралі ДНК, у якій міститься 146 пар основ, закрученого навколо білкової середини (кору), побудованої з восьми молекул гістонів. Нуклеосоми зумовлюють суперкомпактизацію молекул ДНК у цих ділянках. Крім того, електронна мікроскопія виявляє в ядрі структури, які вважають продуктами транскрипційної активності хроматину. До них належать перихроматинові фібрили товщиною 3...5 нм, перихроматинові гранули діаметром 45 нм, інтерхроматинові гранули діаметром 21...25 нм.
Ядерце— це найбільш щільна структура ядра (щільність ядерця в 1,5 раза перевищує щільність ядра), яка добре помітна у живій незабарвленій клітині.
Форма ядерець сферична, розмір 1...5 мкм. Ядерце добре забарвлюється, особливо основними барвниками. Це пов'язано з наявністю у ньому великої кількості РНК,концентрація якої тут у два-вісім разів вища, ніж у ядрі, і в два-три рази перевищує концентрацію у цитоплазмі. Кількість ядерець, як правило, відповідає кількості хромосомних наборів. Тому в диплоїдних клітинах їх буває два на ядро.
Ядерце — це не самостійна організована структура, а похідне хромосом, які містять так звані ядерцеві організатори, що здебільшого розташовані у зонах вторинних перетяжок. Останні являють собою локуси хромосом з найбільш високою концентрацією і активністю синтезу РНК в інтерфазі. Ядерце — це місце утворення рибосомних РНК і самих рибосом. ДНК ядерцевого організатора складається із множинних копій генів рРНК: на кожному з них синтезується попередник рРНК, який у зоні ядерця одягається білком; потім тут утворюються субодиниці рибосом.
Субмікроскопічна будова ядерця характеризується наявністю двох основних структур: гранул діаметром 15...20 нм і фібрил товщиною 6...8 нм. Гранулярний компонент часто розташовується у вигляді нитки, нуклеолонеми товщиною 0,2 мкм. Фібрилярний компонент ядерця — це рибонуклеопротеїнові тяжі, попередники рибосом, а гранули — субодиниці рибосом, що дозрівають. Навколо ядерця знаходиться компактна зона навколо ядерцевого гетерохроматину. Конденсований хроматин також розміщений поміж петлями нуклеолонеми.
Каріоплазма – це рідка частина ядра, в якій містяться ядерні структури, аналог гіалоплазми у цитоплазматичній частині клітини. Після видалення з ядер ДНК, РНК, гістонових та мембранних білків вони не втрачають своєї цілісності, незважаючи на майже повну втрату хроматину і мембран. Під електронним мікроскопом у таких ядрах виявлено комплекси пор разом з фібрилярним периферійним шаром, ядерцеві фібрили та численні фібрили, що лежать у міжхроматинових районах. Весь комплекс цих структур, побудований з негістонових білків, отримав назву білкового ядерного матриксу, який можна вважати аналогом цитоматриксу цитоплазми. До білкового ядерного матриксу входять компоненти ядерної оболонки, ядерця, каріоплазми. Матрикс ядра відіграє важливу роль як у підтриманні загальної структури інтерфазного ядра, так і в процесах його метаболізму.
Репродукція клітин. Розмноження клітин — це одне з найважливіших біологічних явищ і є проявом загальної закономірності, яка полягає у тому, що неодмінною умовою існування біологічних систем протягом досить довгих проміжків часу є їхня репродукція. Розмноження клітин здійснюється шляхом поділу вихідної клітини. Це положення є одним з основних у клітинній теорії.
Клітинний цикл (cyclus cellularis). Весь період існування клітини від поділу до поділу або від поділу до смерті називають клітинним циклом. У дорослому організмі вищих хребетних тварин і людини клітини різних органів і тканин мають різну здатність до поділу і, таким чином, різний клітинний цикл. Поділові клітини передує подвоєння її хромосомного набору, а отже, і кількості ДНК. Це подвоєння відбувається у строго визначеному періоді інтерфази і лише після цього процесу починається поділ клітини. Нещодавно відкрито білок циклін, який регулює вступ клітини у мітоз. Зменшення швидкості синтезу цикліну збільшує тривалість інтерфази.
Весь клітинний цикл поділяють на чотири періоди: власне мітоз (М), пресинтетичний (G1), синтетичний (S) та пост-синтетичний (G2) періоди інтерфази. Позначення пресинтетичного періоду літерою G походить від англійського «grow» — рости. Це період посиленого росту молодої клітини, головним чином за рахунок нагромадження клітинних білків. У цей період починається підготовка клітини до синтезу ДНК, який відбувається в наступному S-періоді. Якщо в експерименті викликати пригнічення синтезу білка або іРНК у G1-періоді, то перехід клітини в S-період блокується. У періоді G1синтезуються ферменти, необхідні для утворення попередників ДНК, метаболізму РНК і білка.
У синтетичному S-періоді (абревіатура S походить від слова «synthesis» – синтез) подвоюється кількість ДНК і відповідно число хромосом. S-період є вузловим у клітинному циклі. Тільки та клітина, яка пройшла цей період, може вступати у мітоз. Рівень синтезу РНКу S-періоді зростає відповідно зі збільшенням кількості ДНК і досягає свого максимуму в G2-періоді. У S-періоді також відбувається подвоєння центріолей клітинного центру.
Постсинтетичний період G2 циклу ще називають премітотичним, оскільки він має велике значення для наступного поділу. У цьому періоді відбувається синтез іРНК, необхідної для проходження мітозу. Крім того, синтезується рРНК рибосом, що визначають поділ клітини. У цей же час синтезуються білки мітотичного веретена — тубуліни. У кінці G2-періоду, або з початком мітозу, синтез РНКрізко падає і повністю зупиняється під час мітозу. Синтез білка під час мітозу також знижується, а потім досягає максимуму в G2-періоді, повторюючи загалом характер синтезу РНК.
Описаний клітинний цикл притаманний клітинам, що зберігають здатність до поділу. Але поряд з цим в організмі є і клітини, які ніби виходять з циклу. Це так звані клітини G0-періоду. Вони не проходять S-періоду і не поділяються, знаходячись у стані спокою. Це клітини, що тимчасово або остаточно перестали поділятися. Існує кілька типів клітин G0-періоду. До першого типу належать стовбурові клітини різних тканин (наприклад, кровотворні). Це малодиференційовані клітини, які, зберігаючи здатність до поділу, на довгий час виходять з циклу, вступаючи в G0-період. До другого типу належать клітини, які, втрачаючи здатність до поділу, спеціалізуються, проходять диференціацію. Серед клітин цього типу можна розрізнити два підтипи. Одні клітини, ставши на шлях диференціації, назавжди втрачають здатність до поділу, деякий час функціонують і потім гинуть. Прикладом таких клітин можуть бути зрілі клітини крові, клітини епідермісу тощо. Клітини другого підтипу після диференціації не втрачають здатності до поділу і, коли потрібно, можуть повертатися у цикл. Наприклад, клітини печінки при видаленні частини органа починають синтезувати ДНК і вступають у мітоз. Третій тип клітин G0-періоду – це високодиференційо-вані клітини, які у дорослому організмі незворотньо втрачають здатність до поділу і мають термін життя, який дорівнює термінові життя цілого організму. Це, наприклад, нервові клітини.
Мітоз (mitosis), каріокінез,або непрямий поділ, є універсальним способом розмноження клітин. Сама назва «мітоз» походить від грецького слова «мітос» — нитка, під яким розуміють нитки хромосом. Під час мітозу внаслідок конденсації еухроматину в ядрі стають видимими вже редупліковані хромосоми, які за допомогою ахроматинового мітотичного апарату розходяться до полюсів клітини, після чого поділяється клітинне тіло. Для зручності вивчення єдиного процесу мітотичного поділу розрізняють чотири фази: профазу, метафазу, анафазу, телофазу (рис. 1.11).
Профаза характерна тим, що хроматиновий малюнок інтерфазного ядра зникає, а натомість з'являються ниткоподібні щільні тільця, які добре забарвлюються і мають назву хромосом. Спочатку вони відокремлені одна від одної не дуже чітко (рання профаза, або стадія щільного клубка), а у кінці профази окремі хромосоми вже добре видно (пізня профаза, або стадія пухкого клубка). Оскільки редуплікація ДНК відбулася в S-періоді інтерфази, то кожна хромосома вже є подвійною структурою. Але в профазі ця подвійність ще не проявляється через щільне прилягання сестринських хромосом (або хроматид) одна до одної. У кінці профази або на початку наступної стадії – метафази – зникає ядерце внаслідок інактивації рибосомних генів у зоні ядерцевих організаторів. Одночасно руйнується ядерна оболонка, яка розпадається на фрагменти, а потім на дрібні мембранні пухирці. Крім того, зменшується кількість елементів гранулярної ендоплазматичної сітки (як цистерн, так і рибосом), що відповідає значній редукції рівня синтезу білка.
Під час профази відбувається ще один дуже важливий для поділу клітини процес — формування веретена поділу внаслідок розходження центріолей до полюсів клітини. До кожного полюсу відходить подвійна центріоля — диплосома. З розходженням диплосом починають формуватися мікротрубочки, які відходять від периферійних ділянок материнської центріолі кожної диплосоми. Сформований апарат поділу в тваринних клітинах має веретеноподібну форму і складається з двох центросфер з центріолями всередині них і волокон веретена, яке лежить між ними. Всі ці три структури побудовані з мікротрубочок, які утворюються внаслідок полімеризації тубулінів у зоні центріолей. Крім того, центрами організації мікротрубочок веретена є спеціальні структури хромосом — кінетохори, розташовані в зонах первинних перетяжок. В результаті у веретені поділу утворюється два типи волокон: центральні, що йдуть від полюсів до центру веретена, і кінетохорні, або хромосомні, які сполучають хромосоми з одним із полюсів клітини і виникають пізніше.
Метафаза починається від того моменту, коли хромосоми, вільно розташовані в цитоплазмі після розчинення ядерної оболонки, починають рухатися до екватору клітини. Цей процес носить назву метакінезу. У середині метафази хромосоми, вишикувавшись в екваторіальній площині веретена, утворюють так звану метафазну пластинку, або материнську зірку, в якій центромерні ділянки хромосом обернені до центру, а їхні плечі — до периферії. У кінці метафази можна побачити, що кожна хромосома складається з двох сестринських хроматид, плечі яких лежать паралельно. їх розділяє щілина, і вони лишаються з'єднаними лише у ділянці центромери. Метафаза займає третину часу всього мітозу.
Анафаза. Усі сестринські хроматиди одночасно в усіх хромосомах втрачають зв'язок між собою в ділянці центромери і синхронно починають рухатися до протилежних полюсів клітини зі швидкістю 0,2...0,5 мкм/хв. Вони орієнтовані центромерами до полюсів, а плечима — до екватора. Це найкоротша стадія мітозу, яка займає лише кілька процентів від усього часу. Анафаза — дуже важлива стадія мітозу, саме на цій стадії 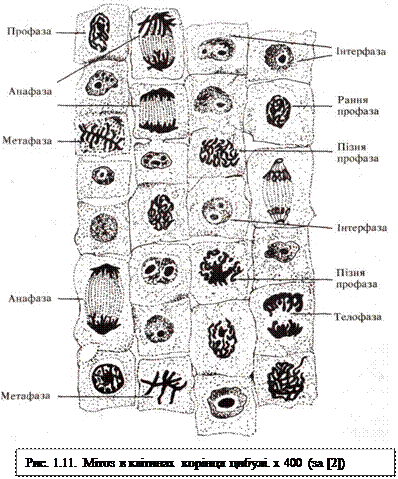 відбувається відокремлення двох ідентичних наборів хромосом та їхнє переміщення до протилежних кінців клітини. Крім руху самих хромосом до полюсів, додатково розходяться ще й самі полюси. Механізм руху хромосом точно невідомий. Останнім часом більшість дослідників дотримується гіпотези «ковзних ниток», згідно з якою сусідні мікротрубочки веретена, взаємодіючи між собою та скоротливими білками, тягнуть хромосоми до полюсів.
відбувається відокремлення двох ідентичних наборів хромосом та їхнє переміщення до протилежних кінців клітини. Крім руху самих хромосом до полюсів, додатково розходяться ще й самі полюси. Механізм руху хромосом точно невідомий. Останнім часом більшість дослідників дотримується гіпотези «ковзних ниток», згідно з якою сусідні мікротрубочки веретена, взаємодіючи між собою та скоротливими білками, тягнуть хромосоми до полюсів.
Телофаза починається із зупинки двох диплоїдних наборів хромосом. Орієнтація хромосом лишається такою ж, як і в анафазі, тобто центромерами до полюса. Вони деконденсуються, збільшуються в об'ємі. У місцях їх контактів з мембранними пухирцями цитоплазми відновлюється ядерна оболонка. Здійснюється формування нових ядерець. У телофазі також відбувається поділ клітинного тіла, що має назву цитотомії, або цитокінезу. У тваринних клітин він проходить шляхом утворення перетяжки у ділянці колишнього екватора: плазмолема вгинається всередину клітини. При цьому в кортикальному шарі цитоплазми в зоні екватора розташовуються циркулярно актинові фібрили. Скорочення такого кільця завершується поділом клітинного тіла. В утворених клітинах починається новий G1-період.
Хромосоми — це щільні паличко- або ниткоподібні тільця діаметром 0,2...2 мкм і довжиною у,людини від 1,5 до 10 мкм, які добре забарвлюються основними барвниками і які помітні в ядрі клітини під час мітотичного поділу (рис. 1.11). Назву їм дав В. Вальдеєр. Поява хромосом — найяскравіша ознака поділу клітини.
Відомо, що хромосоми не зникають із закінченням мітозу, а існують у ядрі і під час інтерфази, але завдяки деконденсації вони набувають іншого вигляду і їх не видно як окремі тільця. В основі як інтерфазних, так і мітотичних хромосом лежать молекули дезоксирибонуклеопротеїнів — ДНП. Останнім часом переважає думка, що кожна хромосома побудована з однієї гігантської молекули ДНП, запакованої у відносно коротке тільце — власне мітотичну хромосому. Знайдено, що довжина індивідуальних лінійних молекул ДНК може досягати сотень мікрометрів і навіть кількох сантиметрів. Серед хромосом людини найбільша перша хромосома. Вона містить ДНК загальною довжиною молекули близько 7 см. Сумарна довжина молекул ДНК у всіх хромосомах однієї клітини людини становить близько 170 см, що відповідає масі 6•10-12 г. Виявлено, що у мітотичній хромосомі гігантська молекула дезоксирибонуклеопротеїну утворює бічні петлі. Типова хромосома людини може містити 2600 петель, кожна з яких утворена ділянкою хроматинової фібрили із середньою довжиною близько 400 нм (0,4 мкм). Компактизація петель веде до утворення таких структур як хромонемні фібрили (хромонеми). Шляхом взаємодії всіх компонентів хромосоми відбувається кінцева компактизація хроматину у вигляді мітотичної хромосоми.
 Морфологію мітотичних хромосом найкраще вивчати у метафазі та на початку анафази, коли вони є найбільш конденсованими. У кожній хромосомі (рис. 1.12) можна відзначити звужене місце — первинну перетяжку (центромеру), яка поділяє хромосому на два плеча. Хромосоми, що мають плечі рівної довжини, називають метацентричними. Якщо одне плече коротше, хромосома має назву субметацентричної. Третій різновид хромосом має центромеру, розташовану майже на кінці. Вона відокремлює коротеньке, часто малопомітне плече — це так звані акроцентричні хромосоми. У ділянці первинної перетяжки розташований кінетохор, який є центром організації мікротрубочок, що утворюють хромосомні нитки веретена поділу. Деякі хромосоми мають також вторинні перетяжки, які локалізовані поблизу від одного з кінців хромосоми і відокремлюють так званий супутник хромосоми. Вторинні перетяжки ще називають ядерцевими організаторами, тому що саме у цих ділянках на початку інтерфази утворюється ядерце. Ділянки хромосом, розташовані поблизу первинної перетяжки, мають назву прицентромерних районів. Кінцеві ділянки плечей називають теломерами.
Морфологію мітотичних хромосом найкраще вивчати у метафазі та на початку анафази, коли вони є найбільш конденсованими. У кожній хромосомі (рис. 1.12) можна відзначити звужене місце — первинну перетяжку (центромеру), яка поділяє хромосому на два плеча. Хромосоми, що мають плечі рівної довжини, називають метацентричними. Якщо одне плече коротше, хромосома має назву субметацентричної. Третій різновид хромосом має центромеру, розташовану майже на кінці. Вона відокремлює коротеньке, часто малопомітне плече — це так звані акроцентричні хромосоми. У ділянці первинної перетяжки розташований кінетохор, який є центром організації мікротрубочок, що утворюють хромосомні нитки веретена поділу. Деякі хромосоми мають також вторинні перетяжки, які локалізовані поблизу від одного з кінців хромосоми і відокремлюють так званий супутник хромосоми. Вторинні перетяжки ще називають ядерцевими організаторами, тому що саме у цих ділянках на початку інтерфази утворюється ядерце. Ділянки хромосом, розташовані поблизу первинної перетяжки, мають назву прицентромерних районів. Кінцеві ділянки плечей називають теломерами.
Кожний вид рослинних і тваринних організмів має специфіку числа, розмірів та будови хромосом. Сукупність цих ознак хромосомного набору називається каріотипом. Каріотип людини характеризує наявність 23 пар хромосом, серед яких 22 пари аутосом і одна пара статевих хромосом. Серед останніх, які носять назву гоносом, розрізняють X- та Y-хромосоми. Хромосоми людини поділяють за розмірами на сім груп — А, В, С, D, Е, F, G. Кількість хромосомних наборів у клітині позначають терміном плоїдністьі літерою п. Соматичні клітини мають подвійний (диплоїдний) набір хромосом (2п), статеві клітини — одинарний гаплоїдний (п) набір. Якщо клітина має 3п набір хромосом, то вона триплоїдна, Якщо 4п — тетраплоїдна тощо. Велика кількість хромосомних наборів позначається терміном поліплоїдія.
Значні успіхи на шляху вивчення морфології хромосом були досягнуті внаслідок застосування спеціальних методів їхнього фарбування —так званого диференційного забарвлення, уперше запропонованого Т. Касперсоном. Виявилось, що хромосоми забарвлюються неоднорідно. Важливим є те, що кожна хромосома при такому диференційованому забарвленні має свій неповторний малюнок. Останній дає змогу чіткої ідентифікації кожної хромосоми в наборі, а також складання так званих хромосомних карт з виясненням локалізації у них певних генів. Зараз встановлена хромосомна локалізація понад 200 генів людини.
Ендомітоз. На основі мітотичного циклу виник ряд механізмів, за допомогою яких кількість спадкового матеріалу може бути збільшена при збереженні сталості числа клітин. Ендомітоз або ендорепродукція — це утворення клітин із збільшеним вмістом ДНК, що відбувається внаслідок блокування мітозу на певних етапах. Зупинка мітозу можлива після G2-періоду, тоді клітина може пройти наступний цикл реплікації ДНК. Це зумовить зростання кількості хромосомних наборів у чотири-вісім і більше разів. Морфологічно такі ядра нічим не відрізняються від ядер з диплоїдним набором, лише збільшеним об'ємом. Зупинка мітозу можлива також у профазі або метафазі, коли порушується функція веретена поділу. Нарешті, можливе проходження клітиною всіх фаз мітозу, включаючи телофазу, без поділу клітинного тіла. Так виникають двох'ядерні клітини (наприклад, клітини печінки у людини). У результаті серії ендомітозів виникають гігантські поліплоїдні клітини червоного кісткового мозку — мегакаріоцити, які можуть мати число хромосомних наборів до 64 п. Необхідно відзначити, що поліплоїдизація зустрічається лише у спеціалізованих диференційованих клітинах.
Мейоз— своєрідна форма клітинної репродукції, яка характерна для процесу утворення статевих клітин. Мейоз включає два послідовних мітотичних поділи, між якими відсутня інтерфаза. У результаті мейозу утворюються клітини з гаплоїдним набором хромосом. Характерною особливістю профази мейозу є кросинговер — обмін гомологічними ділянками хромосом, який є одним із суттєвих факторів мінливості організмів.
Дата добавления: 2015-09-18; просмотров: 3338;