ОРНИТИНОВЫЙ ЦИКЛ
Мочевина - основной конечный продукт азотистого обмена, в составе которого из организма выводится избыток азота.Экскреция мочевины в норме составляет около 25 г/сут. Синтезируется только в печени.
Катаболизм аминокислот и образование аммиака происходят во многих тканях. Длятранспорта азота из тканей в печень используется 3 соединения:глутамин, аланин и аммиак.
Орнитиновый цикл в печени выполняет 2 функции:
• превращение азота аминокислот в мочевину, которая экскретируется ипредотвращает накоплениетоксичных продуктов, главным образомаммиака;
• синтез аргинина и пополнение его фонда в организме.
Полный набор ферментов орнитинового цикла есть только в гепатоцитах(рис.6).
В последней реакции цикла, которая происходит только в печени, под действием аргиназыаргинин распадается на мочевину и орнитин. Орнитин вновь поступает в митохондрии и повторно включается в цикл синтеза мочевины.
Молекула мочевины содержит 2 атома азота:
• первый атом N* поступает в цикл в виде аммиака, образующегося в митохондриях. Реакция катализируется митохондриальной карба-моилфосфатсинтетазой I.
• второй атом N** вводится в мочевину из аспарагиновой кислоты.
Аммиак, используемый карбамоилфосфатсинтетазой I,поставляется в печень главным образом из кишечникас кровью воротной вены. Роль других источников, в том числе окислительного дезаминирования глутаминовой кислоты, существенно меньше.
Аспарагиновая кислота, необходимая для синтеза аргининосукцината,образуется в печени с использованиемаминогруппы аланина, который поступает главным образомиз мышц и клеток кишечника. Oбразующийся в орнитиновом цикле фумарат в результате 2 реакций превращается в оксалоацетат, из которого путем трансаминирования образуется аспартат. Таким образом, с орнитиновым циклом сопряженцикл регенерации аспартата из фумарата. Пируват, образующийся в этом цикле из аланина, используется для глюконеогенеза.
Еще одинисточник аспартата для орнитинови цикла - трансаминирование глутамата с оксалоацетатом.
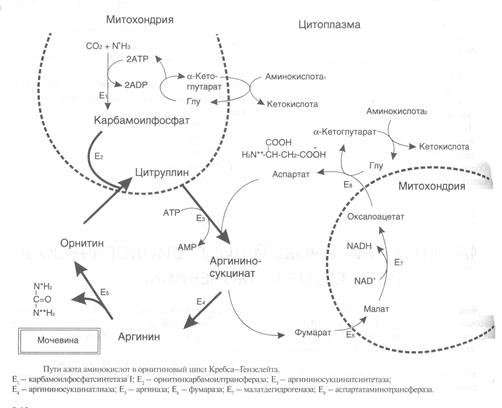
Рисунок 6
В орнитиновом цикле расходуются 4 макроэргические связи 3 молекул АТР на каждый оборот цикла. Однако процесс превращения аминокислот в безазотистые остатки и мочевинуобеспечивает сам себяэнергией:
- прирегенерации аспартата из фумарата на стадии дегидрирования малатаобразуется NADH, который может обеспечить синтез 3 макроэргических связей (рис.7);
- приокислительном дезаминироваиии глутамата в разных органах такжеобразуется NADH, который может обеспечить образование 3 макроэргичес-ких связей.
Затратыэнергии происходят ипри трансмембранном переносевеществ, связанном с синтезом и экскрециеймочевины. Первые 2 реакции орнитинового цикла исходят в митохондриях, а последующие 3 - в цитоплазме.Цитруллин, образующийся в митохондрии,должен бьпъ перенесен в цитозоль, а орнитин, образующийся цитозоле,должен быть перенесен в митохондрию. Кроме того, в почкахперенос мочевины из крови в мочу происходит за счет градиента ионов натрия, создаваемого К+, Na+- АТРазой.
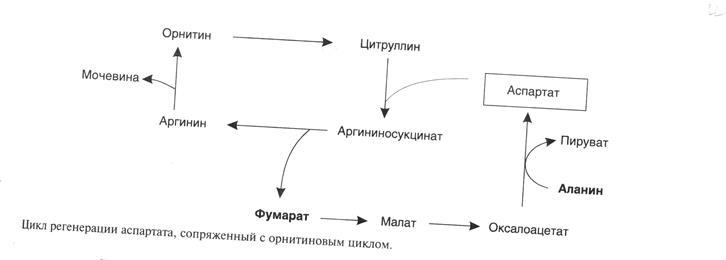
Рисунок 7
Отдельные ферменты орнитинового цикла обуживаются не только в печени, но и в других тканях. В энтероцитах, например, имеется 2 первых фермента и, следовательно, может синтезироваться цитруллин. В почках обнаружены 3-й и 4-й ферменты цикла. Цитруллин, образовавшийся в энгероцитах, может поступать в почки и превращаться там в аргинин, который переносится в печень и гидролизуется аргиназой. Активность этих водящихся в разных органах ферментов значимо ниже, чем в печени.
Дата добавления: 2015-09-18; просмотров: 1661;