ГЕНЕТИЧЕСКИЙ ГРУЗ
Процесс видообразования с участием такого фактора, как естественный отбор, создает разнообразие живых форм, приспособленных к условиям обитания. Среди разных генотипов, возникающих в каждом поколении благодаря резерву наследственной изменчивости и перекомбинации аллелей, лишь ограниченное число обусловливает максимальную приспособленность к конкретной среде. Можно предположить, что дифференциальное воспроизведение этих генотипов в конце приведет к тому, что генофонды популяций будут представлены лишь «удачными» аллелями и их комбинациями. В итоге произойдет затухание наследственной изменчивости и повышение уровня гомозиготности генотипов.
В природных популяциях, однако, наблюдается противоположное состояние. Большинство организмов являются высокогетерозиготными. Отдельные особи гетерозиготны частично по разным локусам, что повышает суммарную гетерозиготность популяции. Так, методом электрофореза на 126 особях рачка Euphausia superba, представляющего главную пищу китов в антарктических водах, изучали 36 локусов, кодирующих первичную структуру ряда ферментов. По 15 локусам изменчивость отсутствовала. По 21 локусу имелось по 3—4 аллеля. В целом в этой популяции рачков 58% локусов были гетерозиготными и имели по 2 аллеля и более. В среднем у каждой особи по 5,8% гетерозиготных локусов. Средний уровень гетерозиготности у растений составляет 17%, беспозвоночных — 13,4, позвоночных — 6,6%. У человека этот показатель равен 6,7%. Столь высокий уровень гетерозиготности нельзя объяснить только мутациями в силу относительной их редкости.
Наличие в популяции нескольких равновесно сосуществующих генотипов в концентрации, превышающей по наиболее редкой форме 1%1, называют полиморфизмом. Наследственный полиморфизм создается мутациями и комбинативной изменчивостью. Он поддерживается естественным отбором и бывает адаптационным (переходным) и гетерозиготным (балансированным).
Адаптационный полиморфизм возникает, если в различных, но закономерно изменяющихся условиях жизни отбор благоприятствует разным генотипам. Так, в популяциях двухточечных божьих коровок Adalia bipunctata при уходе на зимовку преобладают черные жуки, а весной—красные (рис. 11.7). Это происходит потому, что красные формы лучше переносят холод, а черные интенсивнее размножаются в летний период.
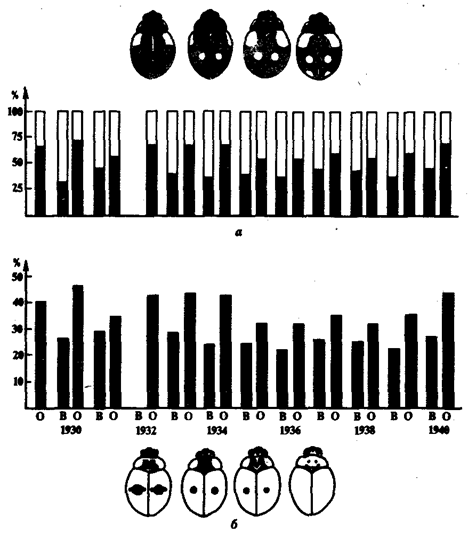
Рис. 11.7. Адаптационный полиморфизм у двухточечных божьих коровок:
а—соотношение черной (зачернено) и красной форм при весеннем (В) и осеннем (О) сборе; б—частота доминантного аллеля черной окраски в весенней и осенней популяциях
Балансированный полиморфизм возникает, если отбор благоприятствует гетерозиготам в сравнении с рецессивными и доминантными гомозиготами. Так, в опытной численно равновесной популяции плодовых мух Drosophila melanogaster, содержащей поначалу много мутантов с более темными телами (рецессивная мутация ebony), концентрация последних быстро падала, пока не стабилизировалась на уровне 10% (рис. 11.8). Анализ показал, что в созданных условиях гомозиготы по мутации ebony и гомозиготы по аллелю дикого типа менее жизнеспособны, чем гетерозиготные мухи. Это и создает состояние устойчивого полиморфизма по соответствующему локусу.
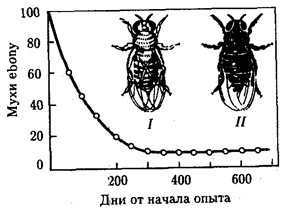
Рис. 11.8. Балансированный полиморфизм по локусу окраски тела в опытной популяции плодовых мух: I—серая муха (дикий тип), II—мутантная муха с черной окраской тела
Явление селективного преимущества гетерозигот называют сверхдоминантностью. Механизм положительного отбора гетерозигот различен. Правилом является зависимость интенсивности отбора от частоты, с которой встречается соответствующий фенотип (генотип). Так, рыбы, птицы, млекопитающие предпочитают обычные фенотипические формы добычи, «не замечая» редких.
В качестве примера рассмотрим результаты наблюдений, выполненных на обыкновенной наземной улитке Cepaea nemoralis, раковина у которой бывает желтая, различных оттенков коричневого цвета, розовая, оранжевая или красная. На раковине может быть до пяти темных полос. При этом коричневая окраска доминирует над розовой, а они обе — над желтой. Полосатость является рецессивным признаком. Улитки поедаются дроздами, использующими камень как наковальню, чтобы разбить раковину и добраться до тела моллюска. Подсчет числа раковин разной окраски вокруг таких «наковален» показал, что на траве или лесной подстилке, фон которых достаточно однороден, добычей птиц чаще оказывались улитки с розовой и полосатой раковиной. На пастбищах с грубыми травами или в живых изгородях с более пестрым фоном чаще поедались улитки, раковины которых окрашены в светлые тона и не имели полос.
Самцы относительно редких генотипов могут иметь повышенную конкурентоспособность за самок. Селективное преимущество гетерозигот обусловливается также явлением гетерозиса. Повышенная жизнеспособность межлинейных гибридов отражает, по-видимому, результат взаимодействия аллельных и неаллельных генов в системе генотипов в условиях гетерозиготности по многим локусам. Гетерозис наблюдается в отсутствие фенотипического проявления рецессивных аллелей. Это сохраняет скрытыми от естественного отбора неблагоприятные и даже летальные рецессивные мутации.
В силу разнообразия факторов среды обитания естественный отбор действует одновременно по многим направлениям. При этом конечный результат зависит от соотношения интенсивности разных векторов отбора. Конечный результат естественного отбора в популяции зависит от наложения многих векторов отборов и контротборов. Благодаря этому достигается одновременно и стабилизация генофонда, и поддержание наследственного разноообразия.
Балансированный полиморфизм придает популяции ряд ценных свойств, что определяет его биологическое значение. Генетически разнородная популяция осваивает более широкий спектр условий жизни, используя среду обитания более полно. В ее генофонде накапливается больший объем резервной наследственной изменчивости. В результате она приобретает эволюционную гибкость и может, изменяясь в том или ином направлении, компенсировать колебания среды в ходе исторического развития.
В генетически полиморфной популяции из поколения в поколение рождаются организмы генотипов, приспособленность которых неодинакова. В каждый момент времени жизнеспособность такой популяции ниже уровня, который был бы достигнут при наличии в ней лишь наиболее «удачных» генотипов. Величину, на которую приспособленность реальной популяции отличается от приспособленности идеальной популяции из «лучших» генотипов, возможных при данном генофонде, называют генетическим грузом. Он является своеобразной платой за экологическую и эволюционную гибкость. Генетический груз — неизбежное следствие генетического полиморфизма.
Дата добавления: 2015-09-11; просмотров: 1272;