Лекция №2. Эволюция насекомых и близких к ним групп.
Членистоногие произошли от каких-то древних многощетинковых кольчатых червей.
У этих червей предротовая лопасть слилась с первыми двумя сегментами тела, образовав акрон. На акроне расположены усики, первые – соответствующие конечностям второго сегмента и верхняя губа, расположенная перед ротовым отверстием, которая образовалась путем укорочения и слияния первой пары ног кольчатых червей.
Главным новшеством членистоногих стал наружный скелет, образовавшийся благодаря уплотнению покровов. В панцирь в виде щитков, разделенных перепонками, оделись сегменты тела, ноги стали суставчатыми, состоящими из уплотненных колец, соединенных мягкими подвижными участками покровов. Как и у кольчатых червей, ноги вблизи основания несут жабры, но у членистоногих конечности стали еще выполнять третью роль – роль челюстей благодаря появлению жевательного выступа на внутренних поверхностях их сближенных оснований.
Первоначальное строение у членистоногих было простое: большое число одинаковых сегментов с ногами, спереди располагался акрон, сзади тельсон – хвостовая лопасть. Из яйца вылуплялась личинка (сходная с трохофорой) сразу с 4 первичными сегментами. Дальнейшее развитие шло за счет добавления сегментов между хвостовой лопастью и задним личиночным сегментом.
Жесткий наружный скелет препятствует росту, поэтому потребовались линьки, во время которых старый покров отстает, лопается и сбрасывается, отделяясь от лежащего под ним пока еще нежного нового. Новый сначала растягивается, а затем затвердевает в более крупных формах. Так у членистоногих ступенчато, от линьки к линьке происходит рост.
Членистоногие возникли в воде как придонные организмы и их первичная эволюция шла в водной среде.
Наружный скелет открыл большие возможности усложнения и усовершенствования конструкции тела членистоногих, позволил повысить их двигательную активность и создал возможность усложнения движений тела и его конечностей. Совершенствование конструкции тела членистоногих первоначально шло в сторону разделения функций отдельных участков тела и оформления их в своеобразно устроенные отделы, называемые тагмами. Передние сегменты вместе с акроном объединились в головной отдел, в котором сосредотачивались функции ориентации в пространстве и захвата и первичной обработки пищи, средний отдел брал на себя задачи передвижения с помощью ног, задний отдел сохранял и усиливал функцию пищеварения и размножения. Первоначально отделы обосабливались нерезко, что позволяло им частично менять свой сегментный состав, так некоторые сегменты груди переходили к голове, некоторые сегменты брюшка присоединялись к груди, также могло происходить объединение и разделение отделов.
Древнейшие ископаемые членистоногие, жившие примерно 570 миллионов лет назад, относятся к трилобитам и ракообразным. Трилобиты – группа членистоногих, наиболее сходная с их общим предком и вымершая 260 млн. лет назад. От них происходят хелицеровые и в более раннее время – общие предки ракообразных и трахейных.
У трилобитов обособлены от прочих и слиты с акроном в головной щит четыре первых несущих ноги сегмента. Ноги этих сегментов совершенно одинаковые и такие же, как на лежащих позади них свободных туловищных сегментах. Все ноги снабжены жабрами и челюстными выступами. Трилобиты вели придонный образ жизни, питаясь разлагающимися остатками водорослей и мелкими донными организмами. Пищу движением ног они продвигали к ротовому отверстию сзади наперед по средней борозде с помощью челюстных выступов.
Предки хелицеровых перешли к активному роющему поиску более крупной добычи в верхнем слое донного грунта. В связи с этим они утратили усики, которые мешали рытью с помощью переднего края головного щита и не могли служить по прямому назначению, и у них также ослабело зрение.
У хелицеровых ноги стали отличаться друг от друга: передние потеряли жабры, из них первая пара превратилась в хватательные (хелицеры) и перестала использоваться при ходьбе, три следующие пары головного щита и две присоединившиеся к ним пары туловищных усилили свою роль в передвижении и начальной переработке пищи. Следующая за хелицерами пара конечностей затем все больше приобретала роль щупалец и утрачивала значение при ходьбе и захвате пищи, она получила название ногощупальцы (педипальпы) – так была восполнена утрата усиков. Передний отдел хелицеровых, совмещающий роль головы и груди, получил название головогрудь. Ноги заднего отдела тела (брюшка) напротив перестали использоваться при ходьбе и стали главным образом органами дыхания, преобразовывались в копулятивные органы или стали использоваться для выделения паутины. Задняя часть брюшка вообще лишилась ног.
От примитивных хелицеровых идут две линии эволюции.
Эволюция первого направления происходила в водной среде. Она дала мечехвостов, ракоскорпионов и затем скорпионов, которые в дальнейшем вышли на сушу, превратив жаберные ножки в легочные мешки. Для этого направления характерно быстрое завершение формирования головогрудного отдела с полным включением двух бывших сегментов туловища в единую головогрудь.
Второе направление эволюции привело к формированию настоящих паукообразных – животных сухопутных, не имеющих прямого родства со скорпионами. В этой линии долго сохранялось самостоятельное существование двух сегментов с ходильными ногами, присоединившихся функционально к головогрудному отделу. Оно завершилось их полным слиянием с первичной головогрудью.
Выход паукообразных на сушу сопровождался превращением жабр в легкие.
При этом у паукообразных, как и у скорпионов, жабры погружались в глубь тела в особый карман, предохранявший дыхательную поверхность от высыхания. При дальнейшей эволюции легкие замещались или дополнялись трахеями. Так, у пауков трахеи заместили сначала задние легочные мешки, а затем и передние. Некоторые мелкие паукообразные не имеют ни легких, ни трахей и дышат всей поверхностью тела. При переходе таких групп в более сухие местообитания у них могут возникать трахеи как замена не легочного, а кожного дыхания.
Эволюция в пределах паукообразных (за исключением клещей) шла в основном по путям выработки различных вариантов хищного образа жизни.
Клещи среди паукообразных занимают особое место. Это не единая по происхождению группа, а по меньшей мере две разных, совершенно независимо перешедших к новым типам питания и в новую среду обитания благодаря сильному уменьшению размеров тела и перестройке комплекса хелицер и педипальп в хоботок. Одна группа перешла к потреблению мертвой растительной пищи и далее к питанию на живых растениях и животных, другая от хищничества перешла к паразитизму и кровососанию.
Другая ветвь, также ведущая свое происхождение от примитивных трилобитов, привела к возникновению ракообразных и трахейных.
У предка этих двух групп (в отличие от трилобитов) усилились преобразования головных конечностей, что было связано с более активным передвижением в поисках пищи и более совершенной ее предварительной обработкой – захватом и измельчением. Кроме уже имевшихся первой пары усиков из передних ног трилобитов сформировалась вторая пара усиков, также превратившихся в орган осязания. Вторые конечности трилобитов стали главными измельчителями пищи – мандибулами, они укрепились и укоротились. Третья и четвертая пара конечностей также усилили функцию обработки пищи и ослабили двигательную. Все головные конечности потеряли жабры. Туловище оставалось состоящим из однообразных члеников без разделения на грудь и брюшко. Главное, что произошло в отличие от хелицеровых – это разделение функций головы и груди (туловища). Это животное можно было бы назвать очень примитивным раком, но раки отличаются от него делением туловища на грудь и брюшко, причем грудь у раков прикрыта сверху и с боков складкой покровов заднего края головы – карапаксом.
Раки от трилобитов отличаются планктонной личинкой, которая выходит из яйца очень слабо развитой, похожей на трохофору. У нее развиты только две пары усиков и мандибулы, затем идет приращение последующих сегментов и их конечностей. Начало развития личинки с двух сегментов после акрона, а не с 4, как у трилобитов и трахейных, связано с переходом к планктонному существованию. Подвижная планктонная личинка ракообразных (в отличие от донной малоподвижной личинки трилобитов), несмотря на свою уязвимость при нападении хищников, давала преимущество при расселении в новые пригодные для жизни места.
Ракообразные – чрезвычайно разнообразные водные животные. Они прошли очень большой путь эволюционных преобразований в водной среде, заселив все местообитания вплоть до влаги между песчинками на побережьях и до подземных вод. Они в ограниченных пределах выбрались даже на сушу – самая успешная попытка принадлежит мокрицам.
Не менее 400 млн. лет назад от общего ствола с примитивными ракообразными отделилась ветвь, перешедшая к жизни на суше. Эти животные – общие предки многоножек и насекомых – приобрели трахейное дыхание, утратив жабры.
Ранние этапы эволюции трахейных изучены еще очень плохо. Отсутствие связи трахей с предшествовавшими им жабрами, как это проявляется у хелицеровых, говорит о том, что скорее всего вышедшие на сушу предки трахейных перешли сначала на кожное дыхание во влажной среде и лишь затем выработали трахеи. Трахейные вышли на сушу еще до заселения ее растениями, они освоили гниющие береговые выбросы водной растительности. Необходимость передвигаться по тесным скважинам в выбросах, чтобы избежать иссушающего действия воздуха, и более активный подвижный образ жизни сделали ненужным удвоение числа усиков и их вторая пара редуцировалась – сложился характерный тип трахейного животного с одной парой усиков и ногами без жабр. Развитие происходило с нарастанием числа сегментов по мере роста и начиналось с вылупления из яйца шестиногой личинки, на новых сегментах ноги сначала появлялись в виде зачатков, затем со следующими линьками приобретали полное развитие. В некоторых группах трахейных нарастание ног на вновь добавляющихся сегментах затормозилось и прекратилось, и таким образом появились взрослые шестиногие с телом, разделенным на грудной отдел с ногами и брюшной – без ног. Уменьшение числа ног облегчает быстрое передвижение (длинные многоножки ползают медленно). Большинство многоножек питается мертвой мягкой растительной пищей и ротовые конечности у них развиты слабо. Некоторые многоножки впоследствии перешли к хищничеству, которое потребовало усиление ротового аппарата и особенно хватательной функции. У хищных многоножек (класс губоногих) не произошло существенного усиления исходных ротовых органов, а функцию схватывания и обездвиживания жертвы взяли на себя ноги первого туловищного сегмента, превратившиеся в крючки с ядовитыми железами. Изменилось и индивидуальное развитие. В связи с повышением активности личинки у губоногих рождаются сразу многоногими, иногда даже сразу с окончательным числом сегментов. Три других класса многоножек сохраняют исходный тип питания мертвыми растительными остатками или отчасти переходят и к живой растительной пище. Среди нехищных многоножек выделяются два класса – пауроподы и двупарноногие, у которых произошло попарное слияние сегментов со спинной стороны, но сохранилось прежнее число ног – на каждом вторичном сегменте по две пары ног.
Особую группу составляют шестиногие многоножки, которых долгое время считали, а кое-кто и сейчас считает настоящими насекомыми.
Их насчитывается 3 класса, из которых наиболее многочисленны и известны ногохвостки. Ротовые органы в этих трех группах приспособлены к поеданию только сравнительно мягкой пищи, они погружены вглубь головы и этой особенностью они резко отличаются от настоящих насекомых и сходны с настоящими многоножками.
Самые примитивные первичнобескрылые настоящие насекомые – щетинохвостки.
Они сохранили сравнительно крепкие и хорошо развитые ротовые придатки, хотя также питаются мертвой растительной пищей, но более грубой и жесткой. У щетинохвосток из частей недоразвитых брюшных ножек 8-9 сегментов брюшка формируется яйцеклад, характерный и для крылатых насекомых.
Величайшим событием в истории жизни на Земле было приобретение насекомыми способности к полету.
Произошло это 300-320 миллионов лет назад, когда насекомые вроде щетинохвосток заселили кроны деревьев. Предпосылкой к полету стали прыжки с ветки на ветку или вниз во спасение от нападающих хищников. При этом преимущество получали те формы, у которых была выше способность к планированию. Для этого служили боковые складки – лопасти ceгментов, особенно грудных. Такие складки были уже у трилобитов и они первоначально служили защитой от механических повреждений нежных частей тела и защитой от нападения сверху. У насекомых особое развитие получили лопасти среднего и заднего сегментов груди. Когда они стали длиннее, их положение стало зависеть от гибких движений грудных сегментов при работе ног – при напряжении мышц, соединяющих спинку сегмента с основаниями ног, лопасти приподнимались. Это открыло возможность управлять планированием, а затем складывать лопасти вдоль тела и взмахивать ими. Для активного опускания крыльев послужили продольные мышцы, выгибающие вверх спинные участки. Складывание крыльев сделало возможным их значительное удлинение и превратило их в конце концов в орган активного полета. Так для насекомых открылись огромные эволюционные возможности.
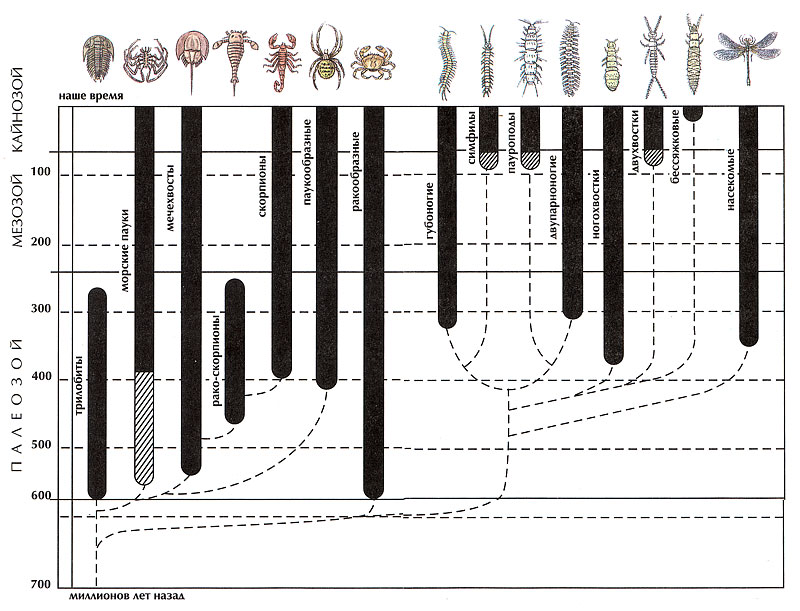
Родственные связи и пути происхождения крупных групп членистоногих.
| <== предыдущая лекция | | | следующая лекция ==> |
| Показатели затрат и себестоимости продукции | | | Векторы и линейные операции гад векторами |
Дата добавления: 2015-11-10; просмотров: 1582;