Синтез пуриновых нуклеотидов
Пуриновые основания, образующиеся в процессе переваривания нуклеиновых кислот в кишечнике, практически не используются, поэтому их синтез осуществляется из низкомолекулярных предшественников, продуктов обмена углеводов и белков.
Пуриновый цикл собирается постепенно путем присоединения необходимых фрагментов к рибозо-5-фосфату. Рибозо-5-фосфат при участии АТФ и пирофосфокиназы превращается в 5-фосфорибозил-1-пирофосфат (ФРПФ).
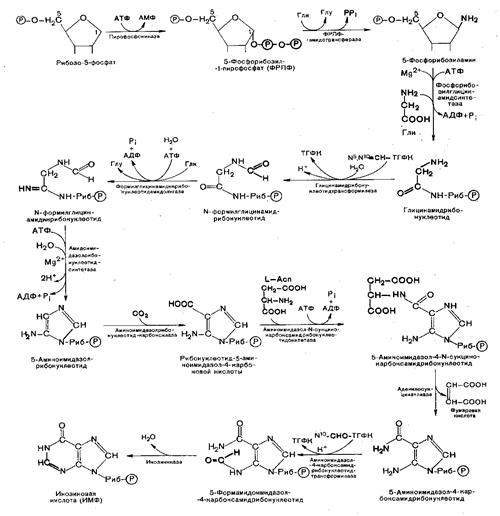
Затем осуществляется 9 последовательных реакций, завершающися сборкой первого нуклеотида – инозиновой кислоты (ИМФ) с гипоксантином в качестве основания. ИМФ является предшественником пуриновых нуклеотидов.
ФРПФ взаимодействует с глутамином, являющимся донором NH2-группы, в результате чего образуется b-5-фосфорибозил-амин. На следующей стадии присоединяется вся молекула глицина к свободной NH2-группе b-5-фосфорибозил-амина с образованием глицинамидрибонуклеотида. Затем цепь удлиняется за счет присоединения формильной группы из N5, N10-метил-ТГФК (ТГФК – тетрагидрофолиевая кислота) с образованием формилглицинамидрибонуклеотида. На формильную группу формилглицинамидрибонуклеотида переносится далее амидная группа глутамина и синтезируется формилглицинамидинрибонуклеотид. На следующей стадии замыкается пятичленное имидазольное кольцо и образуется 5-аминоимидазолриӯбонуклеотид, который способен акцептировать СО2 с образованием рибонуклеотида 5-аминоимидазол-4-карбоновой кислоты. В последующем двухступенчатом процессе, в котором участвуют аспарагиновая кислота и АТФ, образуется 5-аминоимидазол-4 карбоксамидрибонуклеотид и освобождается фумаровая кислота. В этих реакциях азот аспарагиновой кислоты включается в 1-е положение будущего пуринового ядра. Последний углеродный атом пиримидинового остатка кольца пурина вводится в виде формильного остатка (источник N10-метил-ТГФК), который присоединяется к 5-NH2-группе. После этого отщепляется молекула воды и второе кольцо замыкается. В результате образуется первый пуриновый нуклеотид – ИМФ. АМФ и ГМФ образуются из ИМФ.
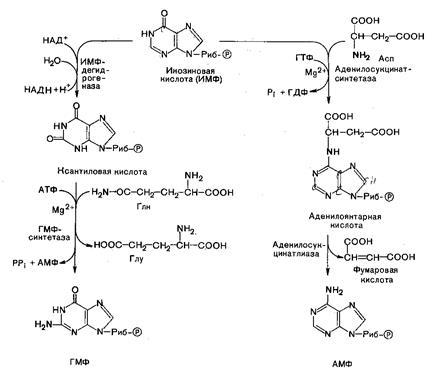
В реакциях участвуют по два фермента. Образование ГМФ из ИМФ катализирует ИМФ-дегидрогеназа и ГМФ-синтеза, а образование АМФ из того же предшественника катализируется последовательным действием аденилосукцинатсинтетазы и аденилосукцинат-лиазы.
В синтезе АМФ из ИМФ специфическое участие принимают аспарагиновая кислота, являющаяся донором NH2-группы, и ГТФ в качестве источника энергии; промежуточным продуктом реакции является аденилоянтарная кислота. Биосинтез ГМФ, напротив, начинается с дегидрогеназной реакции ИМФ с образованием ксантозиловой кислоты.
Превращение АМФ и ГМФ в соответствующие нуклеозидди- и нуклеозидтрифосфаты также протекает в 2 стадии при участии специфических нуклеозидмонофосфат- и нуклеозиддифосфаткиназ:
ГМФ + АТФ  ГДФ + АДФ
ГДФ + АДФ
ГДФ + АТФ ГТФ + АДФ
Пуриновые основания у млекопитающих активно синтезируются в печени. Клетки головного мозга, эритроциты, нейтрофилы особенно зависят от поступления пуринов. Синтез нуклеотидов, используя готовые пуриновые основания, осуществляется только в эмбриональной, регенерирующей, опухолевой ткани. Все ферменты синтеза и распада пуриновых нуклеотидов находятся в цитозоле клеток.
Синтез пуриновых нуклеотидов тормозится по принципу обратной связи, т.е. ингибированием первой стадии переноса аминогруппы глутамина на ФРПФ. Фермент, катализирующий эту стадию, является аллостерическим регуляторным ферментом. Особенность механизма регуляции заключается в том, что избыток ГМФ в клетках оказывает аллостерическое торможение только на свой собственный синтез, не влияя на синтез АМФ, и, наоборот, накопление АМФ подавляет свой синтез, не ингибируя синтеза ГМФ.
Дата добавления: 2015-07-06; просмотров: 4710;