Механизм действия гормонов пептидной природы
Рецепторы пептидных гормонов локализованы на плазматических мембранах и имеют три домена – внеклеточный, внутримембранный и цитоплазматический. При присоединении гормона к рецептору развиваются процессы, приводящие к образованию мессенджеров. Мессенджеры активируют ферменты-протеинкиназы, являющиеся АТФ-фосфотрансферазами. Они катализируют фосфорилирование белков-регуляторов путем переноса фосфатных групп от АТФ к гидроксильным группам остатков серина, треонина, тирозина в составе этих белков.
В систему вторичных мессенджеров входят циклические АМФ (цАМФ) и ГТФ (цГТФ), ионы кальция, ионозилтрифосфат, диацилглицерол и др.
Цикло-АМФ выступает в роли вторичного посредника при действии таких гормонов, как ЛГ, ФСГ, АКТГ, липотропины, МСГ, антидиуретический гормон (АДГ), тиреолиберин, паратин, кальцитонин, глюкагон, секретин, катехоламины, мелатонин, гастрин, гистамин.
цАМФ образуется из АТФ под действием аденилатциклазы, встроенной в цитоплазматическую мембрану клеток. Аденилатциклаза – липопротеид, имеющий регуляторную и каталитическую субъединицы. Регуляторная единица связана с гормональными рецепторами через ГТФ-связывающие, или G-белки, которые выполняют роль универсальных сопрягающих белков. Установлено два типа сопрягающих белков: Gs-активирующие белки, которые стимулируют активность аденилатциклазы и увеличивают продукцию цАМФ, и ингибирующие Gi-белки, которые подавляют активность аденилатциклазы и снижают продукцию цАМФ. Оба белка состоят из 3-х субъединц – a, b и g. При взаимодействии гормона со специфическим рецептором клетки происходят конформационные изменения G-белка. В результате освобождается a-субчастица, которая взаимодействует с ГТФ, и этот комплекс активирует аденилатциклазу. G-белки обладают ГТФ-азной активностью, и поэтому в последующем происходит расщепление ГТФ с образованием ГДФ. Комплекс G-белок-ГТФ оказывает на аденилатциклазу ингибирующее действие, и передача гормонального сигнала завершается (рис.18).
цАМФ взаимодействует с ферментом протеинкиназой, который путем фосфорилирования активирует в клетках определенные белки – ферменты и обеспечивает физиологические эффекты данного гормона.
Протеинкиназы могут существовать в двух формах: неактивной, состоящей из 4-субъединиц, и активной.
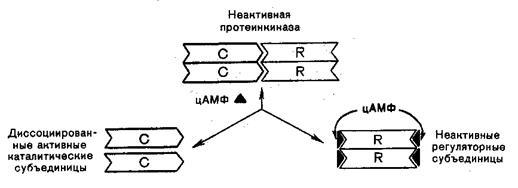
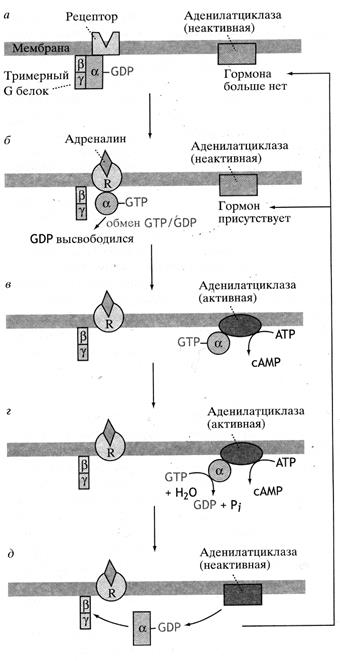
Рис.18. Регуляция активности аденилатциклазы адреналином
В отсутствие цАМФ фермент представляет комплекс из двух каталитических (С) и двух регуляторных R субъединиц. При наличии цАМФ комплекс активируется путем обратимой диссоциации на одну регуляторную и две свободные каталитические субъединицы. Каталитические субъединицы с участием АТФ фосфорилируют другие регуляторные белки. Дефосфорилирование белков и возвращение в исходное функциональное состояние осуществляется с помощью фермента фосфопротеидфосфофатазы по схеме:
Фосфопротеид + Н2О ® белок + Н3РО4
Инактивация цАМФ в клетке происходит под действием фермента фосфодиэстеразы.
Наряду с цАМФ в клетках обнаружен и цГМФ, который обнаружен в гуанилатциклазной реакции. Через гуанилатциклазную систему реализуются эффекты атрипептидов, фактора роста, гистамина, глутамата.
Для ряда гормонов и нейротрансмиттеров вторичным посредником являются ионы кальция. К агентам, которые активируют рецептоуправляемые кальциевые каналы, относятся рецепторы вазопрессина, окситоцина, гастрина и т.д. Под действием гормонального стимула рецептор взаимодействует с G-связывающими белками и воздействует на рецептороуправляемые кальциевые каналы, и Cа2+ поступает из внешней среды или внутреннего депо. Уровень кальция в клетке повышается. Дальнейшая передача гормонального стимула Cа2+ происходит в разных клетках двумя путями: непосредственно воздействием на активность белков-ферментов, ионных насосов, селективных каналов проницаемости, сократительных и других белков; специфических протеинкиназ. Этот путь осуществляется взаимодействием Cа2+ с высокочувствительным к кальцию регуляторным белком-кальмодуллином, который обнаруживается во всех ядросодержащих клетках, имеет 4 участка связывания кальция. Вне кальция кальмодуллин находится в неактивном состоянии. Связывая кальций, он активирует и инициирует кальцийзависимые протеинканизы, которые фосфорилируют белки, а последние вызывают конечный эффект гормона.
Вопросы и задачи
1. От каких факторов зависит реализация биологического эффекта гормонов?
2. Охарактеризуйте механизм действия стероидных и пептидных гормонов.
3. Что такое вторичные посредники и какие из них вам известны?
4. Назовите основные эффекты повышения концентрации цАМФ?
5. Какое действие будет оказывать тироксин на клетки в присутсвии ингибиторов транскрипции или трансляции?
Рекомендуемая литература
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия: Учебник.– М.: Медицина, 1998. – 704 с.
2. Жеребцов Н.А., Попова Т.Н., Артюхов В.Г. Биохимия.– Воронеж: Изд-во Воронеж. гос. ун-та, 2002. – 696 с.
3. Камилов Ф.Х., Давлетов Э.Г. Биохимия гормонов и механизм гормональной регуляции обмена веществ. – Уфа: Гилем, 1998. – 268 с.
4. Киршенблат Я.Б. Общая эндокринология: Учеб. пособие для ун-тов.– М.: Высшая школа, 1971. – 384 с.
5. Комов В.П. Биохимия. – М.: Дрофа, 2004. – 640 с.
6. Красильникова Л.А., Авксентьева О.А., Жмурков В.В., Садовниченко Ю.А. Биохимия растений / Под ред. Л.А. Красильниковой. – Ростов н/Д.: «Феникс»; Харьков: Торсинг, 2004. – 224 с.
7. Филлипович Ю.Б. Основы биохимии: Учебник для хим. и биол. спец. ун-тов и ин-тов.– М.: Изд-во «Агар», 1999. – 512 с.
8. Чиркин А.А. Практикум по биохимии: Учеб. пособие. – Мн.: Новое издание, 2002. – 512 с.
9. Эллиот В. Биохимия и молекулярная биология / В. Эллиот, Д. Эллиот; Под ред. А.И. Арчакова, М.П. Кирпичникова, А.Е. Медведева, В.П. Скулачева. – Пер. с англ. О.В. Добрыниной, И.С. Севериной, Е.Д. Скоцеляс и др. – М.: МАИК «Наука / Интерпериодика», 2002. – 446 с.
Часть II.
ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ В ОРГАНИЗМЕ
Дата добавления: 2015-07-06; просмотров: 2784;