Соотношение концепций регуляционизма и стохастизма
В результате дискуссии стало очевидным, что очень важно четко различать факторы определяющие и факторы регулирующие плотность популяции.
Регулирование означает наличие у популяции тенденции снижать численность при превышении определенного уровня и увеличивать, когда этот уровень не достигнут. Другими словами, регулирование популяции, по определению, может быть только результатом одного или более зависящих от плотности процессов, влияющих на рождаемость, смертность, иммиграцию или эмиграцию.
С другой стороны, уровень численности будет определяться всеми факторами и процессами, воздействующими на популяцию, как зависящими так и не зависящими от плотности. Факторами, зависимыми от плотности, называют такие, удельное (т.е. в расчете на одну особь) воздействие которых меняется при изменении плотности популяции, а независимыми от плотности - те, удельное воздействие которых при разном уровне плотности сохраняется одним и тем же.
Например, если в популяции оленей ежегодно погибает от волков 30% всех особей (а абсолютное число погибших соответственно колеблется год от года в зависимости от колебаний численности популяции), то пресс хищников в данном случае выступает в качестве фактора, независимого от плотности. Если же, наоборот, доля погибших от волков оленей будет колебаться, достигая 50% в годы высокой численности и только 5% в годы низкой численности, то пресс хищников будет фактором, зависимым от плотности.
Фактически получается, что и Николсон и Адреварте правы, но отвечали они на разные вопросы. Сторонники стохастизма отвечают на вопрос «Почему в этот год такая численность популяции?», т.к. численность в каждый момент определяется условиями местообитания. Сторонники регуляционизма отвечают на вопрос «Почему изменения численности имеют пределы?». В этом случае мы обнаруживаем механизмы, зависимые от плотности. В частности представления о минимальной и максимальной численности популяции в явной форме содержат указания на зависимость от плотности. Минимальная плотность популяции определяется генетическим процессами и вероятностью встречи партнеров; максимальная численность, в свою очередь, является плотностно-зависимой величиной, определяемой количеством доступных ресурсов.
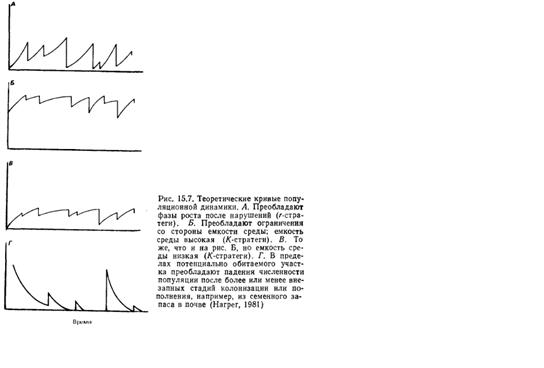
Рис. 6. Теоретические кривые динамики численности популяций: А, Г – популяции повышающие численность после падений; Б, В – популяции «стукающиеся» о верхний предел численности.
Дополнительную аргументацию о непротиворечивости этих объяснений мы находим в работе Мак-Артура и Уилсона «Теория островной биогеографии» (MacArthur, Wilson). Авторы подчеркивают различия в поведении популяций. Некоторые виды (или локальные популяции) большую часть времени тратят на восстановление численности после резких ее спадов или на освоение новых территорий, а другие непрерывно «ударяются» о пределы, накладываемые на нее природными ресурсами, или каким-нибудь иным образом страдают от перенаселенности (проявление сил, зависящих от плотности) (рис. 6). Эти два состояния - два полюса одного континуума. На одном из них наблюдаемая численность популяции обычно отражает (а) уровень последнего снижения численности, (б) время, имевшееся для ее восстановления и (б) присущую популяции в этот период скорость ее увеличения. На другом полюсе численность популяции отражает доступность некого лимитирующего ресурса, сдерживающего дальнейший рост популяции путем ограничения рождаемости, увеличения смертности или стимулирования эмиграции. Полюса этого континуума соответствуют условиям r- и K-отбора Мак-Артура и Уилсона.
Дата добавления: 2015-08-20; просмотров: 950;