Экспрессия рекомбинантных генов в эукариотических системах
При экспрессии эукариотических генов в бактериальных клетках часть рекомбинантных эукариотических белков и некоторые прокариотические белки при высоком уровне биосинтеза переходят в нерастворимое состояние, образуя так называемые тельца включения. Это связано с отсутствием в клетках прокариот систем, обеспечивающих посттрансляциюнные модификации эукариотических белков, происходящие в определенных компартментах эукариотических клеток:
• образование дисульфидных связей (неправильно уложенный белок может оказаться нестабильным и неактивным);
• протеолитическое расщепление предшественника, удаление определенного участка полипептидной цепи с образованием функционального белка;
• гликозилирование: основная модификация, благодаря которой белки приобретают стабильность, или особые свойства;
• модификация аминокислот в составе белка: фосфорилирование, ацетилирование и др.
Все эти модификации структуры белка, а также правильное сворачивание белковой молекулы (фолдинг), которое в клетках эукариотов осуществляют шапероны, влияют на его растворимость, стабильность и биологические функции. Поскольку в бактериальных клетках соответствующие системы посттрансляционных модификаций белков отсутствуют, в них не могут быть получены биологически полноценные рекомбинантные эукариотические полипептиды, что накладывает ограничения на использование бактерий в биотехнологии. Поэтому используют системы эукариотических клеток: дрожжей, насекомых, животных и растений, в геном которых рекомбинантные гены вводятся с помощью трансфекции .
Выбранную эукариотическую систему экспрессии проверяет на наличие всех необходимых условий для требующейся в каждом конкретном случае посттрансляционной модификации и фолдинга продукта вводимого рекомбинантного гена.
Эукариотические экспрессирующие векторы, как и их прокариотические аналоги должны содержать в структуре следующие функциональные модули (рис. 56):
• эукариотический селективный маркер;
• эукариотический промотор;
• соответствующие эукариотические сайты терминации транскрипции и трансляции;
• сигнал полиаденилирования мРНК.
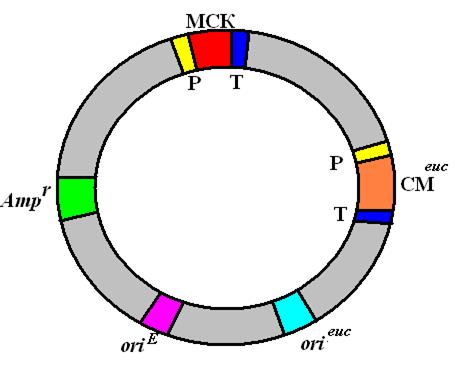
Рис. 56. Обобщенная структура эукариотического экспрессирующего вектора
эукариотический транскриптон с промотором (Р), множественным сайтом клонирования (МСК) и сигналами терминации и полиаденилирования (Т); эукариотический селективный маркер (СМeuk); сайт инициации репликации, функционирующий в клетках эукариот (orieuk); сайт инициации репликации, функционирующий в E. coli (oriЕ); селективный маркер E. coli (Аmрг).
Если вектор функционирует как плазмида, реплицикация которой не зависит от хромосомы, то он должен содержать сайт инициации репликации, функционирующий в хозяйской клетке. Если же вектор предназначен для встраивания в хозяйскую хромосомную ДНК, то для обеспечения рекомбинации он должен нести последовательность, гомологичную определенному участку хромосомной ДНК хозяина (хромосомный сайт интеграции). Поскольку технически многие операции с рекомбинантными ДНК сложнее проводить в клетках эукариот, чем прокариот, большинство эукариотических векторов сконструированы как челночные. Это значит, что эти векторы несут два типа сайтов инициации транскрипции и два типа селективных маркерных генов, одни из которых функционируют в Escherichia coli, a другие — в эукариотических хозяйских клетках. Такие векторные системы экспрессии разработаны для дрожжей, насекомых и клеток млекопитающих.
Экспрессирующие векторы для работы с клетками млекопитающих (ЭВМ)
Внехромосомные экспрессирующие векторы млекопитающих используются для изучения функций и регуляции генов млекопитающих. Кроме того, с их помощью могут быть получены аутентичные рекомбинантные белки, которые потенциально могут быть применимы в медицинских целях для лечения некоторых заболеваний человека. Уже сконструированные экспрессирующие векторы млекопитающих весьма многочисленны, но все они обладают сходными свойствами и похожи на другие эукариотические экспрессирующие векторы (рис. 56).
Полилинкер, представляющий собой множественный сайт клонирования (МСК), и селективный маркер (СМ) находятся под контролем эукариотического промотора и сигнала полиаденилирования или терминации. Обычно используют регуляторные последовательности ДНК вирусов животных (например, цитомегаловируса человека, SV40 или HSV) или генов млекопитающих (гена β-актина, металлотионеина, тимидинкиназы или гена гормона роста). Репликация в клетках E.coli и в клетках млекопитающих обеспечивается соответствующими сайтами инициации репликации: ori плазмиды colEI и, например, обезьяньего вируса 40 (SV40). Для отбора трансформированных клеток E.coli используется ген устойчивости к ампициллину Amp. В качестве селективных маркеров для трансфицированных клеток млекопитающих используют разные гены:
1. Бактериальный ген Neo , кодирующий неомицинфосфотрансферазу, которая обеспечивает устойчивость трансфецированных клеток к токсичному соединению генетицин (G-418).
2. Ген, кодирующий дигидрофолатредуктазу (ДГФР). В этой системе используют клетки с дефектом гена ДГФР, которые не способны расти на среде с метотрексатом. Отобранные клетки пересевают на среды с большей концентрацией метотрексата, отбирая, таким образом, более устойчивые клетки, т.е. содержащие больше копий вектора.
3. Ген фермента глутаминсинтетазы (GS), обеспечивающий устойчивость к цитотоксическому действию метионинсульфоксимина. Вектор, содержащий GS-ген,вводят в культуру клеток млекопитающих и для отбора клеток, несущих большое количество копий вектора, повышают концентрацию метионинсульфоксимина в среде. При этом в хозяйских клетках тоже должна присутствовать GS, поскольку только множественные копии GS-гена могут обеспечивать устойчивость к метионинсульфоксимину.
Дата добавления: 2015-07-14; просмотров: 2849;