ОПЛОДОТВОРЕНИЕ – ТЕОРЕТИЧЕСКИЕ АСПЕКТЫ
Оплодотворение представляет собой слияние двух гамет - ооцита исперматозоида, влекущее за собой слияние их гаплоидных наборов хромосом иразвитие нового организма. Зрелый ооцит к моменту оплодотворения предӯставляет собой одну изнаиболее больших клеток орӯганизма - 110 - 120 мкм в диаметре, окруженнуюблестящей оболочкой (zona pellucida), несколькими слоями клеток лучистого венца(corona radiata) и большим числом клеток яйценосного бугорка (cumuӯlusoophorus). К этому времени в ооците, как правиӯло, уже завершилось первоеделение мейоза, в реӯзультате которого отделилось первое полярное тельӯце, авторое деление мейоза находится на стадии метафазы. Хромосомы располагаются вряд, образуя метафазную пластинку, непосредственно под полярӯным тельцем. Наэтой стадии в ооците происходит блок мейоза, который снимается лишь припроникӯновении сперматозоида. Зрелый сперматозоид имеет уплощенную груӯшевидную головку длиной около 6мкм и шириной в экваториальном сегменте около 4 мкм, состоящую главным образомиз ядра и акросомы, которая соӯдержит литические ферменты и расположена в видешапочки над верхней половиной головки. Хвост сперматозоида имеет длину около 50мкм и начинаӯется от головки в районе шейки, где расположены центриоль икомплекс митохондрий. Митохондрии отвечают за обеспечение энергией процессадвижеӯния сперматозоида, осуществляемого хвостом. В структуре сперматозоида нетничего лишнего, все имеет только одну цель – доставить генетический материал,содержащийся в головке сперматозоида, в ооцит.Способность сперматозоида к оплодотворению in vivo или In vitro появляетсялишь после капацитации, под которой in vivo понимается весь комплексбиохиӯмических и ультраструктурных изменений, которые претерпеваетсперматозоид, проходя путь через женӯский половой тракт до встречи с ооцитом.Эти изменеӯния затрагивают в основном мембрану головки сперӯматозоида.Капацитация in vitro происходит во время обработки и инкубирования спермы дооплодотвореӯния, однако она невозможна при отсутствии в средах для отмывки икультивирования альбумина.При оплодотворении in vivo в трубе присутствует лишь небольшое количествосперматозоидов, продеӯлавших весь долгий путь от влагалища до ампулярногоотдела маточной трубы. Для нормального оплодоӯтворения in vitro их необходимоне менее 50 тыс. на ооцит. Однако при подсчете количества сперматозоиӯдов,непосредственно атакующих ооцит in vitro, их наӯсчитывается до 2 - 3 тыс. Досих пор неясно, почему количество сперматозоидов должно быть так велико вискусственных условиях. Возможно, это объясняется недостаточнойфизиологичностью среды либо больӯшим размером массы cumulus у ооцитов,получаемых при ТВП, чем in vivo.Прежде чем попасть в ооцит, сперматозоид должен преодолеть несколькобарьеров. Первым из них являӯется cumulus, представляющий собой растянутыйматрикс, состоящий преимущественно из полигиалуроновой кислоты, с редкорасположенными клетками. Преодолеть этот барьер может только капацитированныйсперматозоид с интактной акросомой. По мере продвижения внутрь cumulusповедение сперматозоиӯда изменяется: резко возрастает скорость, двумерныедвижения становятся трехмерными - наступает фаза гиперактивности. In vitroпервый барьер сперӯматозоидам помогает преодолевать фермент гиалуронидаза,выделяющаяся при разрушении акросом гибнущих сперматозоидов; однако такаяситуация не является физиологичной.После прохождения cumulus сперматозоид достигаӯет zona pellucida, гдепроисходит его связывание с реӯцептором. В отличие от мышей, рецепторы zоnаpelluӯcida которых давно выделены и охарактеризованы, таковые человека до сихпор точно не определены. Имеется предположение, что это так называемыйанӯтиген к оплодотворению - FA-1, гликопротеин, котоӯрый при добавлении всреду полностью блокирует свяӯзывание сперматозоидов с zona pellucida.Связывание сперматозоида возможно только при интактной акросоме и нормальнойморфологии головӯки, поскольку рецепторы сперматозоида к гопа pelluӯcidaрасположены на акросоме. Затем происходит акросомная реакция - мембранаакросомы и цитоплазматическая мембрана ооцита сливаются, содержимое акросомы(главным образом, фермент акрозин, споӯсобный к локальному растворениюгликопротеинов, из которых состоит гопа pellucida) выбрасывается в местесвязывания с гопа pellucida и сперматозоид инӯтенсивно продвигается сквозьоболочку ооцита, все еще находясь в фазе гиперактивности.Попадая в перивителлиновое пространство, сперӯматозоид прикрепляется крецепторам на мембране ооцита комплементарными рецепторами, расположенӯнымина экваториальной части головки под акросоӯмой. Связывание сперматозоида срецепторами на плазматической мембране мгновенно вызывает так называемуюкортикальную реакцию - массовый экзоцитоз гранул, расположенных по периферииооцита (cortex}, что приводит к необратимым изменеӯниям zona pellucida,делающим ее непроходимой для остальных сперматозоидов. Это является основныммеханизмом предотвращения полиспермии у млекоӯпитающих. Далее начинаетсяпроцесс слияния мембӯран сперматозоида и ооцита, в результате которогосперматозоид целиком как бы заглатывается ооцитом (процесс очень напоминаетфагоцитоз).Сразу после слияния гамет ядерная мембрана сперӯматозоида разрушается,хроматин деконденсируется под влиянием факторов ооплазмы. Одним из этихфакӯторов, возможно, является так называемый фактор деконденсации ядрасперматозоида (SNDF). Ооцит также активируется, мейоз возобновляется,выделяетӯся второе полярное тельце. Механизм активации ооциӯта пока изученнедостаточно, однако известно, что фактор активации выделяетсясперматозоидом. Ядерный материал ооцита окружается оболочкой - формируетсяженский пронуклеус. Вокруг хромаӯтина сперматозоида формируется мужскойпронуклеӯус, в обоих пронуклеусах идет синтез ДНК. Мужской и женскийпронуклеусы начинают движение по наӯправлению друг к другу и встречаются вцентре ооциӯта. Через несколько часов после встречи мембраны пронуклеусовразрушаются, и генетический материал обеих гамет сливается (сингамия). Наэтом этапе проӯцесс оплодотворения завершается - возникает зигота. Хроматинзиготы конденсируется, и хромосомы подгоӯтавливаются к первому делениюдробления.На следующий день после аспирации ооциты переӯносят в лунку со свежейкультуральной средой и очиӯщают от клеток лучистого венца пастеровскойпипетӯкой с оттянутым до диаметра 150 - 160 мкм кончиком и просматривают напредмет присутствия признаков оплодотворения. Пронуклеусы видны в ооците дажепри наблюдении в стереомикроскоп х 80. Они появляӯются, как правило, через 10- 16 ч после добавления сперматозоидов в среду с ооцитами и исчезают через 6- 8 ч после появления. Если присутствуют оба пронуклеуса - оплодотворениесчитается нормальным. Если их не удается обнаружить - оплодотворение несостоялось. Если виден один пронуклеус либо больше двух - произошлоаномальное оплодотворение.Примерно в 30% случаев в ооцитах после оплодоӯтворения in vitro не выявляютсяпронуклеусы, что моӯжет быть связано как с несостоявшейся пенетрацией ооцита(низкая концентрация активных сперматозоиӯдов, дефекты в механизмах адгезиисперматозоида, отсутствие рецепторов на zona pellucida и/или мембраӯнеооцита), так и с незрелостью ооцита на момент опӯлодотворения, а также сналичием хромосомных аноӯмалий у ооцита (диплоидия, анеуплоидия).При наличии в ооплазме одного пронуклеуса (около 3 - 6%) в половине случаевоплодотворение все же происходит, однако пронуклеусы формируются асинӯхронно.Происхождение таких зигот может быть разӯличным: гиногенетическим (изооцита), андрогенетическим (из сперматозоида) либо возникшим в резульӯтатеслияния мужского и женского пронуклеусов. Отличить на практике этоневозможно, поэтому рекоӯмендуется: во-первых, просматривать ооциты наслеӯдующий день после инсеминации несколько раз, чтобы избежать ошибки; во-вторых, не переносить в полость матки эмбрионы, полученные изоднопронуклеарных зигот.Полипронуклеарные зиготы - 3 и более пронуклеӯусов - составляют, как правило,5 - 10% от всех опӯлодотворенных ооцитов и возникают главным образом припроникновении в ооцит более одного сперматозоӯида (в случае добавленияизбыточного количества сперматозоидов при инсеминации, при дефектахкорӯтикальной реакции, при незрелости или перезрелости ооцита). Также описаныслучаи неотделения второго полярного тельца после завершения мейоза,генетичеӯский материал которого формирует третий пронуклеӯус.Полипронуклеарные зиготы, как правило, не развиваются нормально, эмбрионы,полученные из таких зигот нельзя переносить в полость матки. В естественныхусловиях такие эмбрионы иногда имплантируются, однако чаще всего такаябеременность заканчивается выкидышем либо мертворождением.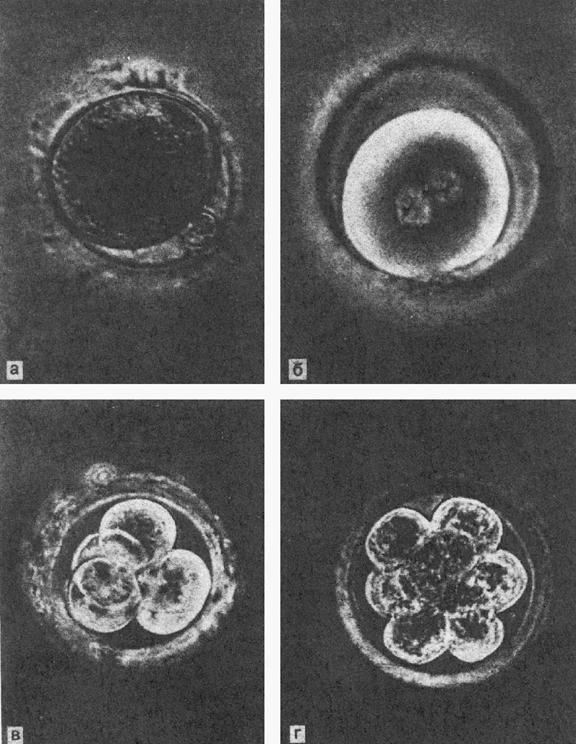 Рис. 4. Яйцеклетки и зародыши человека на различных этапах культивироӯвания.а – ооцит с первым полярным тельцем; б – пронуклеусы: в и г – зародыши настаӯдиях 4 и 8 бластомеров. Микрофотографии живых объектов в фазовомконтрасте. ОЦЕНКА КАЧЕСТВА ЭМБРИОНОВ Эмбрионы, полученные после оплодотворения ооцитов одной и той же пациентки,часто отличаются по скорости дробления и морфологическим параметӯрам.Общепризнанным считается, что максимальную способность к имплантации имеютэмбрионы с наиӯбольшей скоростью дробления, бластомеры которых имеютрегулярную форму, а безъядерные фрагменты отсутствуют. Такие эмбрионы относятк классу 1 (А). Градация эмбрионов по качеству является условной иразличается в разных лабораториях. Однако в осӯнове ее лежит, как правило,характер фрагментации эмбриона, форма и размер бластомеров. Так, эмбрион снеравными бластомерами и/или фрагментами циӯтоплазмы, занимающими менее 10%объема, соответӯствует классу 2 (В); при наличии фрагментации 10 - 50% -классу 3 (С), более 50% - классу 4 (D).Работы по анализу влияния качества переносимых в полость матки эмбрионов начастоту их имплантации многочисленны, но трудносопоставимы в силу разныхметодик оценки качества. Около 20% переносимых эмбрионов А-В классаимплантируются, однако эта частота падает до 1,5% для сильнофрагментированных эмбрионов. Впечатляют результаты клиники Bourn Hall(Англия), в которых показано, что 90% пациенток, забеременевших после ЭКО иПЭ, поӯлучали при переносе эмбрионов хотя бы один эмбрион класса А. В той жеработе не было выявлено зависимоӯсти между качеством переносимых эмбрионов ичастоӯтой невынашивания беременности. СТРАТЕГИЯ ПЕРЕНОСА ЭМБРИОНОВ Количество переносимых эмбрионов обычно соӯставляет не более 2 - 3, посколькупри увеличении чисӯла эмбрионов до 4 и более частота беременности, какправило, не возрастает, но увеличивается риск моногоплодной беременности, чтовлечет за собой серьезӯные проблемы акушерского характера. Однако вотӯдельных случаях допускается перенос большего числа эмбрионов - примногократных неудачных попытках ЭКО и ПЭ и при возрасте пациентки старше 40лет. В данном случае при последующих переносах шанс наступления беременностиснижается, и авторы пытаются использовать возможность наступлениябеӯременности за счет увеличения количества переносиӯмых эмбрионов. Но такойподход не является научӯным, так как в этих случаях не всегда ясна причинанеудач, т. е. мы часто имеем дело с так называемым «бесплодием неяснойэтиологии».Что касается интервала между моментом оплодоӯтворения ооцитов и временемпереноса эмбрионов, то здесь существует несколько основных подходов. На зареразвития метода Эдварде и Стептоу переносили в полость матки эмбрионы,достигшие стадии 8 - 16 блаӯстомеров на 3 - 4-е сутки культивирования,имитируя естественные условия. Однако впоследствии было выявлено, что болееранние эмбрионы также пригодӯны для переноса и имплантируются с той жевероятӯностью.Через 48 ч после аспирации фолликулов (2-е сутки культивирования) эмбрионы,как правило, находятся на стадии 2 - 4 бластомеров, но встречаются и стадии 6- 8 бластомеров. Перенос эмбрионов на 2-е сутки наиӯболее общепринят: ужеимеется возможность отобрать эмбрионы по качеству и скорости дробления,однако пребывание в условиях культуры, являющихся в люӯбом случае менееоптимальными, чем естественные, еще не слишком длительно.Интересен тот факт, что способность эмбриона к имӯплантации напрямую зависитот скорости его дроблеӯния. Так, в исследовании Staessen с соавт. припереносе эмбриона хорошего качества на стадии 2 и 4 бластомеӯров частотаимплантации составила 14 и 21% соответӯственно. В другой работе сообщается,что если перенесенные эмбрионы достигали стадии не более 2 бластомеров,частота наступления беременности составляла 9,3%, но возрастала до 35,8%,если хотя бы один эмбрион был на более чем двуклеточной стаӯдии.На 3-и сутки культивирования нормально развиваӯющиеся эмбрионы достигаютстадии 6 - 8 бластомеров и выше. К этому моменту выбор эмбрионов дляпереӯноса облегчается - часть эмбрионов, на 2-е сутки дроӯбившихся нормально,отстает в развитии либо остаӯнавливается. Однако при анализе качествамногоклеӯточных эмбрионов существует опасность принять безъядерные фрагментыза бластомеры, становящиеся к этой стадии близкими по размеру. В большинствелабораторий, проводящих ЭКО и ПЭ, перенос эмбриоӯнов осуществляется на 2 - 3-и сутки культивирования, причем статистически достоверной разницы междучастотой наступления беременности после переноса на 2-е или 3-и сутки не былообнаружено (соответственно 21,9 и 23,5%). Выбор времени переноса долженосуществляться на основании анализа интенсивности и равномерности дробленияэмбрионов: если выбор эмбрионов на 2-е сутки затруднен, перенос можноотӯложить на 24 ч.На 4 - 5-е сутки пребывания в культуре при примеӯнении стандартных сред иметодик культивирования лишь небольшая часть эмбрионов достигает стадииморулы и бластоцисты, и частота наступления береӯменности не возрастает посравнению с переносом на 2 - 3-и сутки.Однако при использовании культуральных систем, отвечающих метаболическимпотребностям эмбрионов на этой стадии, можно добиться хороших результатов.Так, при культивировании эмбрионов в присутствии клеток линии Vero (клеткипочки обезьяны) было поӯказано, что 60% эмбрионов достигают стадиибластоӯцисты, а перенос бластоцист дает высокий процент беременности.Особенно это относится к пациенткам с повторными неудачами при переносе болееранних эмбрионов. По данным разных авторов, беременность наступала в 37 - 40%случаев переноса эмбрионов на стадии бластоцисты.Серьезным возражением против ко-культивирования эмбрионов человека в присутствииклеток других животных является возможность вирусного заражеӯния, поэтомуактивно разрабатываются среды, оптиӯмизирующие условия культивирования болеепоздних эмбрионов (>3 суток). Примером такой среды может служить средаГарднера (G 2). При культивировании в ней эмбрионов с 3-го по 5-й деньбластоцисты форӯмировались из 52% зигот первого дня, частота имӯплантации ибеременности составляла соответственно 23 и 38%. Сам Гарднер сообщает о 50%-йимӯплантации бластоцист, полученных при последоваӯтельном культивировании всредах G 1,2 (1 - 2-й день) и G 2,2 (3 - 5-й день), в то время как достовернойразӯницы между частотой наступления беременности поӯсле переноса эмбрионов на 3-й и 5-е сутки не наблюӯдалось (66 и 71% соответственно). Возможность культивирования до стадии бластоциӯсты открывает большиевозможности при отборе «лучӯших» эмбрионов, делает более физиологичныммоӯмент попадания эмбрионов в полость матки, а также существенно увеличиваетпроцент имплантации, что позволяет переносить не более 2 бластоцист, неопасаӯясь возникновения осложнений, связанных с многоӯплодием, или снижениявероятности наступления беременности. ТЕХНИКА ПЕРЕНОСА ЭМБРИОНОВ Техника переноса эмбрионов за двадцать лет развития метода практически неизменилась. Здесь мы рассмотрим лишь последовательность действий эмбриолога,не касаясь гинекологических аспектов переноса.Перенос эмбрионов осуществляют через цервикальный канал в полость маткипациентки с помощью специального катетера. Существует большой выбор такихкатетеров, однако наиболее распространенными в мире являются: катетеры Bourn-Wallace, Frydman и Cook-Soft transfer для неосложненных переносов и каӯтетерT.D.T. для осложненных переносов (при загибе матки, извилистом ходе илиспазме цервикального канала).Катетер присоединяют к 1-миллилитровому шприӯцу, набирают столбик (около 0,2мл) свежей, нагретой до 37°С культуральной среды, затем давление со шприцаснимают, шприц переводят в исходное полоӯжение и продолжают набирать: сначалапузырек возӯдуха, затем каплю среды без эмбрионов, опять пузыӯрек воздуха,каплю среды с эмбрионами, отобранныӯми для переноса, пузырек воздуха и ещеодну каплю без эмбрионов. Такая последовательность необходима для обеспечениясохранности эмбрионов во время проӯцедуры переноса и маркирования ихместоположения. Катетер с эмбрионами передают гинекологу для осуӯществленияпереноса: содержимое, за исключением начального столбика среды, всего около20 - 50 мкл, попадает в полость матки. Оставшейся средой промыӯвают катетер исмыв рассматривают под стереомикроскопом, чтобы убедиться, что все эмбрионыперенесеӯны в полость матки. СТИМУЛЯЦИЯ СУПЕРОВУЛЯЦИИ В ПРОГРАММЕ ЭКО И ПЭ Для успешного выполнения программы ЭКО необходимо добиться созреваниянескольких доминантных фоллиӯкулов - суперовуляции. Это значительно повышаетвозӯможность изъятия и оплодотворения яйцеклетки. Кроме того, отмечено, чтопри пересадке нескольких оплодотворенӯных яйцеклеток беременность развиваетсячаще, причем развивается один эмбрион. Этот феномен получил название "функцияпомощи".Период развития от раннего преантрального до преовуляторного фолликула учеловека занимает примерӯно 85 дней, или 3 менструальных цикла (рис.5). После65 дней роста финальная когорта, состояӯщая из 15 - 20 малых полостныхфолликулов, вступает в гонадотропинзависимую фазу роста. В спонтанӯномменструальном цикле в яичнике под влиянием гонадотропинов (Гн), главнымобразом фолликулостимулирующего гормона (ФСГ), развивается один фолӯликул, аостальные подвергаются атрезии. Секреция ФСГ происходит в аденогипофизе ирегулируется гонадотропин-рилизинг гормоном (ГнРГ), вырабатываеӯмымгипоталамусом. Доминантный фолликул обладаӯет высокой стероидогеннойактивностью и продуцируӯет эстрадиол (Е2), необходимый для секреторнойтрансформации эндометрия и обеспечения условий для имплантации эмбриона.Когда секреция E2 достиӯгает критического уровня, происходит резкоевозрастание уровня лютеинизирующего гормона (ЛГ), стиӯмулирующего овуляцию илютеинизацию лопнувшего фолликула. ЛГ, как и ФСГ, секретируется клетӯкамиаденогипофиза и находится под влиянием ГнРГ. Изучение динамики концентрацииE2 в перифериӯческой крови показало, что период активного стероидогенезасоставляет 5 - 6 дней, после чего концентраӯция е2 резко снижается.Мониторниг E2 используется в клинической практике для определения степенифункциональной зрелости доминантного фолликула. На месте разорвавшегосяфолликула формируется желтое тело, которое является основнойстероидпродуцирующей структурой яичника, определяющей изменения концентрацииЕ2 и прогестерона (П) на протяжении лютеиновой фазы менструального цикӯла.Первая беременность, завершившаяся в 1978 г. рождением Луизы Браун, наступилав результате опӯлодотворения in vitro единственного ооцита, аспирированного вспонтанном цикле, и переноса одного эмӯбриона в полость матки. В последующихисследованиӯях была показана низкая эффективность метода при переносе лишьодного эмбриона, которая составляет при работе в естественном цикле не более8 - 15% из расчета на один перенос эмбрионов. Это привело к неӯобходимостииспользования лекарственных препаратов, оказывающих стимулирующее действие нафолликулогенез в яичниках в целях получения нескольӯких преовуляторныхооцитов.Для стимуляции фолликулогенеза и получения неӯскольких преовуляторных ооцитовиспользуются гормональные препараты.
Рис. 4. Яйцеклетки и зародыши человека на различных этапах культивироӯвания.а – ооцит с первым полярным тельцем; б – пронуклеусы: в и г – зародыши настаӯдиях 4 и 8 бластомеров. Микрофотографии живых объектов в фазовомконтрасте. ОЦЕНКА КАЧЕСТВА ЭМБРИОНОВ Эмбрионы, полученные после оплодотворения ооцитов одной и той же пациентки,часто отличаются по скорости дробления и морфологическим параметӯрам.Общепризнанным считается, что максимальную способность к имплантации имеютэмбрионы с наиӯбольшей скоростью дробления, бластомеры которых имеютрегулярную форму, а безъядерные фрагменты отсутствуют. Такие эмбрионы относятк классу 1 (А). Градация эмбрионов по качеству является условной иразличается в разных лабораториях. Однако в осӯнове ее лежит, как правило,характер фрагментации эмбриона, форма и размер бластомеров. Так, эмбрион снеравными бластомерами и/или фрагментами циӯтоплазмы, занимающими менее 10%объема, соответӯствует классу 2 (В); при наличии фрагментации 10 - 50% -классу 3 (С), более 50% - классу 4 (D).Работы по анализу влияния качества переносимых в полость матки эмбрионов начастоту их имплантации многочисленны, но трудносопоставимы в силу разныхметодик оценки качества. Около 20% переносимых эмбрионов А-В классаимплантируются, однако эта частота падает до 1,5% для сильнофрагментированных эмбрионов. Впечатляют результаты клиники Bourn Hall(Англия), в которых показано, что 90% пациенток, забеременевших после ЭКО иПЭ, поӯлучали при переносе эмбрионов хотя бы один эмбрион класса А. В той жеработе не было выявлено зависимоӯсти между качеством переносимых эмбрионов ичастоӯтой невынашивания беременности. СТРАТЕГИЯ ПЕРЕНОСА ЭМБРИОНОВ Количество переносимых эмбрионов обычно соӯставляет не более 2 - 3, посколькупри увеличении чисӯла эмбрионов до 4 и более частота беременности, какправило, не возрастает, но увеличивается риск моногоплодной беременности, чтовлечет за собой серьезӯные проблемы акушерского характера. Однако вотӯдельных случаях допускается перенос большего числа эмбрионов - примногократных неудачных попытках ЭКО и ПЭ и при возрасте пациентки старше 40лет. В данном случае при последующих переносах шанс наступления беременностиснижается, и авторы пытаются использовать возможность наступлениябеӯременности за счет увеличения количества переносиӯмых эмбрионов. Но такойподход не является научӯным, так как в этих случаях не всегда ясна причинанеудач, т. е. мы часто имеем дело с так называемым «бесплодием неяснойэтиологии».Что касается интервала между моментом оплодоӯтворения ооцитов и временемпереноса эмбрионов, то здесь существует несколько основных подходов. На зареразвития метода Эдварде и Стептоу переносили в полость матки эмбрионы,достигшие стадии 8 - 16 блаӯстомеров на 3 - 4-е сутки культивирования,имитируя естественные условия. Однако впоследствии было выявлено, что болееранние эмбрионы также пригодӯны для переноса и имплантируются с той жевероятӯностью.Через 48 ч после аспирации фолликулов (2-е сутки культивирования) эмбрионы,как правило, находятся на стадии 2 - 4 бластомеров, но встречаются и стадии 6- 8 бластомеров. Перенос эмбрионов на 2-е сутки наиӯболее общепринят: ужеимеется возможность отобрать эмбрионы по качеству и скорости дробления,однако пребывание в условиях культуры, являющихся в люӯбом случае менееоптимальными, чем естественные, еще не слишком длительно.Интересен тот факт, что способность эмбриона к имӯплантации напрямую зависитот скорости его дроблеӯния. Так, в исследовании Staessen с соавт. припереносе эмбриона хорошего качества на стадии 2 и 4 бластомеӯров частотаимплантации составила 14 и 21% соответӯственно. В другой работе сообщается,что если перенесенные эмбрионы достигали стадии не более 2 бластомеров,частота наступления беременности составляла 9,3%, но возрастала до 35,8%,если хотя бы один эмбрион был на более чем двуклеточной стаӯдии.На 3-и сутки культивирования нормально развиваӯющиеся эмбрионы достигаютстадии 6 - 8 бластомеров и выше. К этому моменту выбор эмбрионов дляпереӯноса облегчается - часть эмбрионов, на 2-е сутки дроӯбившихся нормально,отстает в развитии либо остаӯнавливается. Однако при анализе качествамногоклеӯточных эмбрионов существует опасность принять безъядерные фрагментыза бластомеры, становящиеся к этой стадии близкими по размеру. В большинствелабораторий, проводящих ЭКО и ПЭ, перенос эмбриоӯнов осуществляется на 2 - 3-и сутки культивирования, причем статистически достоверной разницы междучастотой наступления беременности после переноса на 2-е или 3-и сутки не былообнаружено (соответственно 21,9 и 23,5%). Выбор времени переноса долженосуществляться на основании анализа интенсивности и равномерности дробленияэмбрионов: если выбор эмбрионов на 2-е сутки затруднен, перенос можноотӯложить на 24 ч.На 4 - 5-е сутки пребывания в культуре при примеӯнении стандартных сред иметодик культивирования лишь небольшая часть эмбрионов достигает стадииморулы и бластоцисты, и частота наступления береӯменности не возрастает посравнению с переносом на 2 - 3-и сутки.Однако при использовании культуральных систем, отвечающих метаболическимпотребностям эмбрионов на этой стадии, можно добиться хороших результатов.Так, при культивировании эмбрионов в присутствии клеток линии Vero (клеткипочки обезьяны) было поӯказано, что 60% эмбрионов достигают стадиибластоӯцисты, а перенос бластоцист дает высокий процент беременности.Особенно это относится к пациенткам с повторными неудачами при переносе болееранних эмбрионов. По данным разных авторов, беременность наступала в 37 - 40%случаев переноса эмбрионов на стадии бластоцисты.Серьезным возражением против ко-культивирования эмбрионов человека в присутствииклеток других животных является возможность вирусного заражеӯния, поэтомуактивно разрабатываются среды, оптиӯмизирующие условия культивирования болеепоздних эмбрионов (>3 суток). Примером такой среды может служить средаГарднера (G 2). При культивировании в ней эмбрионов с 3-го по 5-й деньбластоцисты форӯмировались из 52% зигот первого дня, частота имӯплантации ибеременности составляла соответственно 23 и 38%. Сам Гарднер сообщает о 50%-йимӯплантации бластоцист, полученных при последоваӯтельном культивировании всредах G 1,2 (1 - 2-й день) и G 2,2 (3 - 5-й день), в то время как достовернойразӯницы между частотой наступления беременности поӯсле переноса эмбрионов на 3-й и 5-е сутки не наблюӯдалось (66 и 71% соответственно). Возможность культивирования до стадии бластоциӯсты открывает большиевозможности при отборе «лучӯших» эмбрионов, делает более физиологичныммоӯмент попадания эмбрионов в полость матки, а также существенно увеличиваетпроцент имплантации, что позволяет переносить не более 2 бластоцист, неопасаӯясь возникновения осложнений, связанных с многоӯплодием, или снижениявероятности наступления беременности. ТЕХНИКА ПЕРЕНОСА ЭМБРИОНОВ Техника переноса эмбрионов за двадцать лет развития метода практически неизменилась. Здесь мы рассмотрим лишь последовательность действий эмбриолога,не касаясь гинекологических аспектов переноса.Перенос эмбрионов осуществляют через цервикальный канал в полость маткипациентки с помощью специального катетера. Существует большой выбор такихкатетеров, однако наиболее распространенными в мире являются: катетеры Bourn-Wallace, Frydman и Cook-Soft transfer для неосложненных переносов и каӯтетерT.D.T. для осложненных переносов (при загибе матки, извилистом ходе илиспазме цервикального канала).Катетер присоединяют к 1-миллилитровому шприӯцу, набирают столбик (около 0,2мл) свежей, нагретой до 37°С культуральной среды, затем давление со шприцаснимают, шприц переводят в исходное полоӯжение и продолжают набирать: сначалапузырек возӯдуха, затем каплю среды без эмбрионов, опять пузыӯрек воздуха,каплю среды с эмбрионами, отобранныӯми для переноса, пузырек воздуха и ещеодну каплю без эмбрионов. Такая последовательность необходима для обеспечениясохранности эмбрионов во время проӯцедуры переноса и маркирования ихместоположения. Катетер с эмбрионами передают гинекологу для осуӯществленияпереноса: содержимое, за исключением начального столбика среды, всего около20 - 50 мкл, попадает в полость матки. Оставшейся средой промыӯвают катетер исмыв рассматривают под стереомикроскопом, чтобы убедиться, что все эмбрионыперенесеӯны в полость матки. СТИМУЛЯЦИЯ СУПЕРОВУЛЯЦИИ В ПРОГРАММЕ ЭКО И ПЭ Для успешного выполнения программы ЭКО необходимо добиться созреваниянескольких доминантных фоллиӯкулов - суперовуляции. Это значительно повышаетвозӯможность изъятия и оплодотворения яйцеклетки. Кроме того, отмечено, чтопри пересадке нескольких оплодотворенӯных яйцеклеток беременность развиваетсячаще, причем развивается один эмбрион. Этот феномен получил название "функцияпомощи".Период развития от раннего преантрального до преовуляторного фолликула учеловека занимает примерӯно 85 дней, или 3 менструальных цикла (рис.5). После65 дней роста финальная когорта, состояӯщая из 15 - 20 малых полостныхфолликулов, вступает в гонадотропинзависимую фазу роста. В спонтанӯномменструальном цикле в яичнике под влиянием гонадотропинов (Гн), главнымобразом фолликулостимулирующего гормона (ФСГ), развивается один фолӯликул, аостальные подвергаются атрезии. Секреция ФСГ происходит в аденогипофизе ирегулируется гонадотропин-рилизинг гормоном (ГнРГ), вырабатываеӯмымгипоталамусом. Доминантный фолликул обладаӯет высокой стероидогеннойактивностью и продуцируӯет эстрадиол (Е2), необходимый для секреторнойтрансформации эндометрия и обеспечения условий для имплантации эмбриона.Когда секреция E2 достиӯгает критического уровня, происходит резкоевозрастание уровня лютеинизирующего гормона (ЛГ), стиӯмулирующего овуляцию илютеинизацию лопнувшего фолликула. ЛГ, как и ФСГ, секретируется клетӯкамиаденогипофиза и находится под влиянием ГнРГ. Изучение динамики концентрацииE2 в перифериӯческой крови показало, что период активного стероидогенезасоставляет 5 - 6 дней, после чего концентраӯция е2 резко снижается.Мониторниг E2 используется в клинической практике для определения степенифункциональной зрелости доминантного фолликула. На месте разорвавшегосяфолликула формируется желтое тело, которое является основнойстероидпродуцирующей структурой яичника, определяющей изменения концентрацииЕ2 и прогестерона (П) на протяжении лютеиновой фазы менструального цикӯла.Первая беременность, завершившаяся в 1978 г. рождением Луизы Браун, наступилав результате опӯлодотворения in vitro единственного ооцита, аспирированного вспонтанном цикле, и переноса одного эмӯбриона в полость матки. В последующихисследованиӯях была показана низкая эффективность метода при переносе лишьодного эмбриона, которая составляет при работе в естественном цикле не более8 - 15% из расчета на один перенос эмбрионов. Это привело к неӯобходимостииспользования лекарственных препаратов, оказывающих стимулирующее действие нафолликулогенез в яичниках в целях получения нескольӯких преовуляторныхооцитов.Для стимуляции фолликулогенеза и получения неӯскольких преовуляторных ооцитовиспользуются гормональные препараты.  Рис. 5. Цикл развития доминантного фолликула Стимуляция суперовуляции в современных условиях основана на:§ стимуляции выделения собственных эндогенных гонадотропинов кломифеном;§ стимуляции экзогенными гонадотропинами (препараты из мочименопаузальных женщин - препараты человеческого менопаузального гонадотропина(чМГ), содержащие ЛГ и ФСГ (пергонал, неопергонал, хумегон) или ФСГ(метродин);§ стимуляции экзогенными гонадотропинами на фоне блоӯкады собственныхгонадотропинов препаратами агонистов РГ ЛГ (декапептиды - трипторелин,бусерелин, госерелин, нафарелин и др.). Все эти препараты - синтеӯтическиеаналоги гонадотропных рилизинг гормонов, и число их постоянно растет. Все онив 50 - 100 раз активӯнее эндогенных рилизингов. Действие их основано наблокаде собственных гонадотропинов, что позволило некоторым авторам называтьэтот метод «стимуляция суӯперовуляции на фоне гипофизэктомии», хотя на самомделе подавляется секреция только гонадотропинов, а не всех тропныхгипофизарных горӯмонов. На фоне снижения уровня собственных гонадотропиноввведение экзогенных гонадотропинов позволяет регулировать рост доминантныхфолликулов и созреваӯние яйцеклеток.В повседневной практике используются так называемые «длинная схема» и«короткая схема» введения препаратов.При длинной схеме введение агонистов начинают в конце фолликулярной фазыпредыдущего цикла, под контролем снижения ЛГ в крови. После наступленияустойчивого низӯкого «плато» ЛГ приступают к введению гонадотропныхпрепаратов. При короткой схеме агонисты вводятся с 1-го дня менструальногоцикла, гонадотропины - с 3-го или 5-го дня цикла.При любом методе стимуляции суперовуляции постоянно контролируются числофолликулов, темпы их роста и велиӯчина фолликулов. До начала стимуляциисуперовуляции следует проӯводить предварительное гормональное обследование, ав период стимуляции - ультразвуковой (УЗ) и гормоӯнальный мониторинг. Какправило, исследуется уровень эстрадиола крови и толщина эндометрия.Критериями соӯзревания яйцеклетки, точнее говоря, определением времени,оптимального для забора ооцитов, являются: концентрация эстрадиола не менее350 пг/мл на 1 фолликул диаметром более 15 мм, толщина эндометрия 0,8 - 1,0см.Механизм овуляции, в естественном спонтанном менструальном циклеобеспечивающийся выбросом эндогенного ЛГ, в циклах стимуляции суперовуляцииимитируется введением ЛГ-подобного препарата - хорионического гонадотропина(ХГ), получаемого из мочи беременных женщин.Через 35 - 36 ч после введения овуляторной дозы ХГ (10000 ед.) производитсятрансвагинальная пункция яичников (ТВП) в целях аспираӯции зрелых ооцитов.После их оплодотворения in vitro и инкубации в специальной питательной средечерез 48 - 72 ч, в зависимости от интенсивности дробления, производитсяперенос эмбрионов на ранней стадии дробления в полость матки пациентки.После пересадки эмбрионов рекомендуется вводить поддерживающие дозыхорионического гонадотропина по 1500 ед. и дексаметазона по 0,25 мг,последӯний подавляет иммунную реакцию отторжения плодного яйца. Препаратывводят до констатации беременности или до срока очередных менструаций (вслучае неудачи). ОСНОВНЫЕ ПРОБЛЕМЫ, ВОЗНИКАЮЩИЕ ПРИ ПРИМЕНЕНИИ ЭКО И ПЭ Программа ЭКО и ПЭ сложна в связи с тем, что она многоэтапная, но не всеэтапы можно объективно проконтролировать. Остановимся на некоторыхпроблеӯмах, возникающих при применении этого метода.У женщины вовремя одного естественного менструӯального цикла можно получить всреднем не более одной яйцеклетки. Манипулируя с яйцеклеткой, ее легкопотерять или травмировать, поскольку ее размеӯры, включая corona radiata, непревышают 500 мк. Яйцеклетка может быть потеряна, например, в пипетӯке, вкапле питательной среды и т. п. Следовательно, необходим резерв клеток.В связи с этим возникла идея стимуляции так называемой суперовуляции, т. е.получения большего числа яйцеклеток (5 - 10 шт.) за счет примененияанӯтагонистов эстрогенов и менопаузальных гонадотропинов ФСГ и ЛГ. Однакоувеличеӯние количества яйцеклеток влечет за собой другой отӯрицательныйфактор - некоторые из них могут окаӯзаться неполноценными, с нарушениемоогенеза или с хромосомной патологией, а другие могут стать на путь обратногоразвития в процессе фолликулогенеза. Так что не все полученные яйцеклеткимогут быть оплодотворены и нормально развиваться.Высокая частота самопроизвольных абортов (26,2%) после ЭКО являетсяотносительным показателем возможӯной патологии плодов. Эти данныеподтверждаются реӯзультатами исследования хромосом яйцеклеток и эмбриоӯновранних стадий развития. Изӯвестно, что от 35 до 50% нефертилизованных ооцитовв программе ЭКО имеют хромосомные аномалии. Выявлена высокая степеньзависимости хромосомных аномалий от возраста беременных женщин. Так,анэуплоидия диагностируется у 47% женщин после 35, лет и у 25% молодыхженщин; после кломифена - у 55%, поӯсле ЧМГ - у 23%, после чистого ФСГ - у22%. Значиӯтельно чаще хромосомная патология обнаруживается уфрагментированных эмбрионов (33%), чем у нормальӯных (20%).Анализ ооцитов и эмбрионов, полученных в программе ЭКО, свидетельствует отом, что основным фактором, обусловливающим высокую частоту хромосомныханомаӯлий, является возраст беременных. Увеличение частоӯты хромосомнойпатологии с увеличением возраста имеет тенденцию, аналогичную таковой впопуляции, однако показатели этой патологии в 2 раза выше, чем популяционныеданные спонтанной фертильности. Высокая чаӯстота хромосомных аномалий внефертилизованных ооцитах свидетельствует о том, что множественная наведеннаястимуляцией овуляция, или суперовуляция, сопровождается высокой частотойхромосомной патоӯлогии ооцитов, которые созревают в аномальных услоӯвиях вотсутствие физиологической селекции и борьбы за существование доминирующегофолликула, который подавляет рост других фолликулов в одном пуле, возӯможно,не имеющих тенденции к нормальному развиӯтию.Начато внедрение в практику методики флуорисцентной (in situ) гибридизации(FISH), обеспечивающей преимплантационную диагностику врожденной хромосомнойпатологии. Отдельные бластомеры извлекаются из эмбриона и проводится их FISH-анализ. Эмбрионы, по которым получено положительное заключение, могут бытьперенесены в полость матки для дальнейшего развития, а эмбрионы собнаруженной патологией не переносятся (в целях профилактики такихзаболеваний как гемофилия, синдром Патау, Дауна, Эдвардса, Клайнфельтера,моносомия Шершевского-Тернера).Достижением самых последних лет является генная терапия, примеӯнение которойвозможно и на самых ранних стадиях развития эмбрионов. При этом выполняетсяне только диагностика, но и лечение больных зародышей.Особенностью ЭКО является очень высокая частота многоплодных беременностей.Если при естественном зачатии рождается одна двойня на 70 - 80 родов, однатройня на 9000 родов и одна четверня на 50 000 родов, то после ЭКОмногоплодие, включая двойни, тройни и четверни, встречается примерно вполовине(!) всех беременностей. Хорошо известно, что многоӯплодие, особеннокогда беременность больше, чем двумя плодами, создает высокий риск осложненийи для матери, и для ребенка, как в процессе береӯменности, так и родов.Сегодня разработаны способы удаления (редукции) "лишних" плодов (больше двух)под ультразвуковым контролем. На самом деле лишние плоды не удаляют, а путемвведения специальных растворов добиваются того, что они перестают развиватьсяи постепенно рассасываӯются. Эта процедура производится на 7 - 8-й неделебеременности и, как правило, не сопровождается угрозой выкидыша остальныхплодов. Рассасыӯвание лишних плодов может происходить и само по себе, безкаких-либо вмешательств, также до 7 - 8 нед.Имеются данные о возможности возникновения y человека таких «ошибокоплодотворения», как гино- и андрогенез.В программе ЭКО и ПЭ, к сожалению, присутствует возможность возникновенияряда грозных осложнеӯний акушерско-гинекологического характера, которыесуществуют и в естественных условиях. Основное из них - внематочнаябеременность, которая может наӯступить, даже если маточные трубы у женщинынепроӯходимы, а также в случае, если они удалены недостаӯточно радикально(беременность наступает в культях труб). Вероятность наступления внематочнойбеременности в программе ЭКО и ПЭ 10,6%.Следует также знать, что нередко при лечении бесплодия методом ЭКО можетиметь место такое осложнение как синдром гиперстимуляции яичников (СГЯ).Синдром гиперстимуляции яичников (СГЯ) - компӯлекс патологических симптомов,возникающих на фоне приӯменения стимуляторов овуляции, характеризующийсязнаӯчительным увеличением яичников, иногда разрывом их и кровотечением;наличием выпота в брюшной и плевральной полостях, возникновениемтромбоэмболий магистральных сосудов, многоплодной беременностью и пр.В последние годы, в связи с широким распространением лечения бесплодияметодом экстракорпорального оплодотвоӯрения, включающего как первый этапстимуляцию суперӯовуляции, СГЯ привлекает все большее внимание. Первыеописания СГЯ появились еще в 40-х годах, S. Ridberg сообӯщил о значительномувеличении яичников после применеӯния гонадотропинов. В 1962 г. A. Southan,N. Ivanovsky опубликовали случаи СГЯ после приема значительных доз кломифена:в 1970 г. описаны случаи СГЯ при комбинированной терапии кломифеном игонадотропинами [Roӯland M.].Е. Raban и соавт. в 1967 г. предложили классификацию СГЯ, которой пользуютсядо сих пор все клиницисты:• легкая форма,• средней тяжести,• тяжелая.Данные о частоте СГЯ приводит Н. Li в 1993 г. в сборниӯке, посвященномусловиям репродуктивной медицины:— частота легкой формы достигает 23%,— средней тяжести - 10%,— тяжелой - 2%.J. Shenker, D. Weinstein расширили и детализировали классификацию, выделив вкаждой из трех форм еще две степени. Легкая форма:1-я степень - клиническая симптоматика отсутствует, соӯдержаниеэстрадиола в плазме более 150 мкг, в моче прегнадиола выше 10 мг;2-я степень- к этим биохимическим изменениям присоедиӯняется увеличение яичников до 5см в диаӯметре. Форма средней тяжести:3-я степень - боли, чувство тяжести внизу живота и измеӯнения,описанные во 2-й степени;4-я степень - присоединяется тошнота, рвота, понос, размеӯрыяичников - более 5 см в диаметре. Тяжелая форма:5-я степень - к описанным симптомам присоединяется асцит,гидроторакс, яичники более 12 см в диаметре;6-я степень - состояние крайне тяжелое, помимо асцита и гидротораксаразвивается гиперкоагуляция, уменьшается перфузия почек, осложняющаяӯсяолигурией и почечной недостаточностью, яичники резко увеличены, отмечают ихразӯрывы и перекрут.Суммируя представленные данные, можно сказать, что, по-видимому, используемыев программах вспомогательӯной репродукции способы гормональной стимуляциифолликулогенеза и овуляции вносят свой вклад в увеӯличение числа такихошибок. Гибель зародышей в процессе имплантации может быть обусловленапереӯчисленными выше причинами, недостаточностью втоӯрой фазы цикла(изначально существовавшей или возӯникшей в процессе овариальной стимуляции),а такӯже, возможно, особенностями становления иммунологических отношенийэмбриона с организмом матери, вследствие отсутствия трубного периодаразвития, в который эти отношения начинают активно устанавлиӯваться («факторранней беременности» и т.п.).Трудно оценить влияние на гаметогенез и оплодотвоӯрение чисто внешних(экологических, антропогенных, социокультурных и т. п.) факторов. Однако они,безусӯловно, также вносят свой вклад в патологию раннего развития.Таким образом, в большинстве случаев пренатальные потери и приводящая к нимпатология у эмбриоӯнов при использовании методов вспомогательной реӯпродукциивозникают не необходимо и не случайно, а под влиянием определенных факторов,в том числе и приводящих к нарушению регулирующих репродукӯтивный процессмеханизмов. Элиминация же аноӯмальных гамет, зигот и эмбрионов - процесс какнеӯобходимый, так и закономерный, направленный на сохранение постоянствагенотипа популяции. ЗАКЛЮЧЕНИЕ Проблема бесплодия возникла отнюдь не сегодня, она сопровождает человечествоеще с древних времен. Современная медицина достаточно глубоко изучила причинывозникновения женского и мужского бесӯплодия. Разработаны как лекарственные,т.е. консерӯвативные, методы терапии бесплодия, так и оперативӯные.Однако неудовлетворенность достигнутыми резульӯтатами привела к разработкенового метода лечения бесплодия - ЭКО и ПЭ, который быстро завоевал поӯзициюлидирующего в этом направлении.В данной работе представлены основные проблемы программы ЭКО и ПЭ и возможныеосложнения, возникающие на различных ее этапах.Хотя применение метода ЭКО и ПЭ не позволяет в целом решить возникшую встране критическую деӯмографическую ситуацию, тем не менее широкое внедрениеего в практику здравоохранения поможет избавиться от бесплодия сотням тысячсупружеских пар, а следовательно, осуществить также их психолоӯгическуюреабилитацию.В заключение необходимо отметить, что благодаря успехам программы ЭКОдостигнуты положительные реӯзультаты в фундаментальных исследованияхчеловеӯческих гамет и эмбрионов. ЛИТЕРАТУРА 1. Экстракорпоральное оплодотворение и его новые направления в леченииженского бесплодия (теоретические и практические подходы): Руководство дляврачей / Под. ред. В.И.Кулакова, Б.В.Леонова – М., 2001, 782 с.2. Никитин А.И., Китаев Э.М., Савицкий Г.А., Иванова Р.Д., КалашниковаЕ.П. и Устинкина Т.И. Экстракорпоральное оплодотворение у человека споследующей имплантацией эмбриона и рождением ребенка. Арх. анатомии,гистологии и эмбриологии. – Л., 1987, Т.93, вып.10, с.39-43.3. Аншина М.Б., Здановский В.М. Если вам нужен ребенок. – М., 1998, 32с.4. Пшеничникова Т.Я. Бесплодие в браке. – М., 1991, 320с.5. Сметник В.П.. Тумилович Л.Г. Неоперативная гинекология. Руководстводля врачей. – М. 1999, с.476-480.
Рис. 5. Цикл развития доминантного фолликула Стимуляция суперовуляции в современных условиях основана на:§ стимуляции выделения собственных эндогенных гонадотропинов кломифеном;§ стимуляции экзогенными гонадотропинами (препараты из мочименопаузальных женщин - препараты человеческого менопаузального гонадотропина(чМГ), содержащие ЛГ и ФСГ (пергонал, неопергонал, хумегон) или ФСГ(метродин);§ стимуляции экзогенными гонадотропинами на фоне блоӯкады собственныхгонадотропинов препаратами агонистов РГ ЛГ (декапептиды - трипторелин,бусерелин, госерелин, нафарелин и др.). Все эти препараты - синтеӯтическиеаналоги гонадотропных рилизинг гормонов, и число их постоянно растет. Все онив 50 - 100 раз активӯнее эндогенных рилизингов. Действие их основано наблокаде собственных гонадотропинов, что позволило некоторым авторам называтьэтот метод «стимуляция суӯперовуляции на фоне гипофизэктомии», хотя на самомделе подавляется секреция только гонадотропинов, а не всех тропныхгипофизарных горӯмонов. На фоне снижения уровня собственных гонадотропиноввведение экзогенных гонадотропинов позволяет регулировать рост доминантныхфолликулов и созреваӯние яйцеклеток.В повседневной практике используются так называемые «длинная схема» и«короткая схема» введения препаратов.При длинной схеме введение агонистов начинают в конце фолликулярной фазыпредыдущего цикла, под контролем снижения ЛГ в крови. После наступленияустойчивого низӯкого «плато» ЛГ приступают к введению гонадотропныхпрепаратов. При короткой схеме агонисты вводятся с 1-го дня менструальногоцикла, гонадотропины - с 3-го или 5-го дня цикла.При любом методе стимуляции суперовуляции постоянно контролируются числофолликулов, темпы их роста и велиӯчина фолликулов. До начала стимуляциисуперовуляции следует проӯводить предварительное гормональное обследование, ав период стимуляции - ультразвуковой (УЗ) и гормоӯнальный мониторинг. Какправило, исследуется уровень эстрадиола крови и толщина эндометрия.Критериями соӯзревания яйцеклетки, точнее говоря, определением времени,оптимального для забора ооцитов, являются: концентрация эстрадиола не менее350 пг/мл на 1 фолликул диаметром более 15 мм, толщина эндометрия 0,8 - 1,0см.Механизм овуляции, в естественном спонтанном менструальном циклеобеспечивающийся выбросом эндогенного ЛГ, в циклах стимуляции суперовуляцииимитируется введением ЛГ-подобного препарата - хорионического гонадотропина(ХГ), получаемого из мочи беременных женщин.Через 35 - 36 ч после введения овуляторной дозы ХГ (10000 ед.) производитсятрансвагинальная пункция яичников (ТВП) в целях аспираӯции зрелых ооцитов.После их оплодотворения in vitro и инкубации в специальной питательной средечерез 48 - 72 ч, в зависимости от интенсивности дробления, производитсяперенос эмбрионов на ранней стадии дробления в полость матки пациентки.После пересадки эмбрионов рекомендуется вводить поддерживающие дозыхорионического гонадотропина по 1500 ед. и дексаметазона по 0,25 мг,последӯний подавляет иммунную реакцию отторжения плодного яйца. Препаратывводят до констатации беременности или до срока очередных менструаций (вслучае неудачи). ОСНОВНЫЕ ПРОБЛЕМЫ, ВОЗНИКАЮЩИЕ ПРИ ПРИМЕНЕНИИ ЭКО И ПЭ Программа ЭКО и ПЭ сложна в связи с тем, что она многоэтапная, но не всеэтапы можно объективно проконтролировать. Остановимся на некоторыхпроблеӯмах, возникающих при применении этого метода.У женщины вовремя одного естественного менструӯального цикла можно получить всреднем не более одной яйцеклетки. Манипулируя с яйцеклеткой, ее легкопотерять или травмировать, поскольку ее размеӯры, включая corona radiata, непревышают 500 мк. Яйцеклетка может быть потеряна, например, в пипетӯке, вкапле питательной среды и т. п. Следовательно, необходим резерв клеток.В связи с этим возникла идея стимуляции так называемой суперовуляции, т. е.получения большего числа яйцеклеток (5 - 10 шт.) за счет примененияанӯтагонистов эстрогенов и менопаузальных гонадотропинов ФСГ и ЛГ. Однакоувеличеӯние количества яйцеклеток влечет за собой другой отӯрицательныйфактор - некоторые из них могут окаӯзаться неполноценными, с нарушениемоогенеза или с хромосомной патологией, а другие могут стать на путь обратногоразвития в процессе фолликулогенеза. Так что не все полученные яйцеклеткимогут быть оплодотворены и нормально развиваться.Высокая частота самопроизвольных абортов (26,2%) после ЭКО являетсяотносительным показателем возможӯной патологии плодов. Эти данныеподтверждаются реӯзультатами исследования хромосом яйцеклеток и эмбриоӯновранних стадий развития. Изӯвестно, что от 35 до 50% нефертилизованных ооцитовв программе ЭКО имеют хромосомные аномалии. Выявлена высокая степеньзависимости хромосомных аномалий от возраста беременных женщин. Так,анэуплоидия диагностируется у 47% женщин после 35, лет и у 25% молодыхженщин; после кломифена - у 55%, поӯсле ЧМГ - у 23%, после чистого ФСГ - у22%. Значиӯтельно чаще хромосомная патология обнаруживается уфрагментированных эмбрионов (33%), чем у нормальӯных (20%).Анализ ооцитов и эмбрионов, полученных в программе ЭКО, свидетельствует отом, что основным фактором, обусловливающим высокую частоту хромосомныханомаӯлий, является возраст беременных. Увеличение частоӯты хромосомнойпатологии с увеличением возраста имеет тенденцию, аналогичную таковой впопуляции, однако показатели этой патологии в 2 раза выше, чем популяционныеданные спонтанной фертильности. Высокая чаӯстота хромосомных аномалий внефертилизованных ооцитах свидетельствует о том, что множественная наведеннаястимуляцией овуляция, или суперовуляция, сопровождается высокой частотойхромосомной патоӯлогии ооцитов, которые созревают в аномальных услоӯвиях вотсутствие физиологической селекции и борьбы за существование доминирующегофолликула, который подавляет рост других фолликулов в одном пуле, возӯможно,не имеющих тенденции к нормальному развиӯтию.Начато внедрение в практику методики флуорисцентной (in situ) гибридизации(FISH), обеспечивающей преимплантационную диагностику врожденной хромосомнойпатологии. Отдельные бластомеры извлекаются из эмбриона и проводится их FISH-анализ. Эмбрионы, по которым получено положительное заключение, могут бытьперенесены в полость матки для дальнейшего развития, а эмбрионы собнаруженной патологией не переносятся (в целях профилактики такихзаболеваний как гемофилия, синдром Патау, Дауна, Эдвардса, Клайнфельтера,моносомия Шершевского-Тернера).Достижением самых последних лет является генная терапия, примеӯнение которойвозможно и на самых ранних стадиях развития эмбрионов. При этом выполняетсяне только диагностика, но и лечение больных зародышей.Особенностью ЭКО является очень высокая частота многоплодных беременностей.Если при естественном зачатии рождается одна двойня на 70 - 80 родов, однатройня на 9000 родов и одна четверня на 50 000 родов, то после ЭКОмногоплодие, включая двойни, тройни и четверни, встречается примерно вполовине(!) всех беременностей. Хорошо известно, что многоӯплодие, особеннокогда беременность больше, чем двумя плодами, создает высокий риск осложненийи для матери, и для ребенка, как в процессе береӯменности, так и родов.Сегодня разработаны способы удаления (редукции) "лишних" плодов (больше двух)под ультразвуковым контролем. На самом деле лишние плоды не удаляют, а путемвведения специальных растворов добиваются того, что они перестают развиватьсяи постепенно рассасываӯются. Эта процедура производится на 7 - 8-й неделебеременности и, как правило, не сопровождается угрозой выкидыша остальныхплодов. Рассасыӯвание лишних плодов может происходить и само по себе, безкаких-либо вмешательств, также до 7 - 8 нед.Имеются данные о возможности возникновения y человека таких «ошибокоплодотворения», как гино- и андрогенез.В программе ЭКО и ПЭ, к сожалению, присутствует возможность возникновенияряда грозных осложнеӯний акушерско-гинекологического характера, которыесуществуют и в естественных условиях. Основное из них - внематочнаябеременность, которая может наӯступить, даже если маточные трубы у женщинынепроӯходимы, а также в случае, если они удалены недостаӯточно радикально(беременность наступает в культях труб). Вероятность наступления внематочнойбеременности в программе ЭКО и ПЭ 10,6%.Следует также знать, что нередко при лечении бесплодия методом ЭКО можетиметь место такое осложнение как синдром гиперстимуляции яичников (СГЯ).Синдром гиперстимуляции яичников (СГЯ) - компӯлекс патологических симптомов,возникающих на фоне приӯменения стимуляторов овуляции, характеризующийсязнаӯчительным увеличением яичников, иногда разрывом их и кровотечением;наличием выпота в брюшной и плевральной полостях, возникновениемтромбоэмболий магистральных сосудов, многоплодной беременностью и пр.В последние годы, в связи с широким распространением лечения бесплодияметодом экстракорпорального оплодотвоӯрения, включающего как первый этапстимуляцию суперӯовуляции, СГЯ привлекает все большее внимание. Первыеописания СГЯ появились еще в 40-х годах, S. Ridberg сообӯщил о значительномувеличении яичников после применеӯния гонадотропинов. В 1962 г. A. Southan,N. Ivanovsky опубликовали случаи СГЯ после приема значительных доз кломифена:в 1970 г. описаны случаи СГЯ при комбинированной терапии кломифеном игонадотропинами [Roӯland M.].Е. Raban и соавт. в 1967 г. предложили классификацию СГЯ, которой пользуютсядо сих пор все клиницисты:• легкая форма,• средней тяжести,• тяжелая.Данные о частоте СГЯ приводит Н. Li в 1993 г. в сборниӯке, посвященномусловиям репродуктивной медицины:— частота легкой формы достигает 23%,— средней тяжести - 10%,— тяжелой - 2%.J. Shenker, D. Weinstein расширили и детализировали классификацию, выделив вкаждой из трех форм еще две степени. Легкая форма:1-я степень - клиническая симптоматика отсутствует, соӯдержаниеэстрадиола в плазме более 150 мкг, в моче прегнадиола выше 10 мг;2-я степень- к этим биохимическим изменениям присоедиӯняется увеличение яичников до 5см в диаӯметре. Форма средней тяжести:3-я степень - боли, чувство тяжести внизу живота и измеӯнения,описанные во 2-й степени;4-я степень - присоединяется тошнота, рвота, понос, размеӯрыяичников - более 5 см в диаметре. Тяжелая форма:5-я степень - к описанным симптомам присоединяется асцит,гидроторакс, яичники более 12 см в диаметре;6-я степень - состояние крайне тяжелое, помимо асцита и гидротораксаразвивается гиперкоагуляция, уменьшается перфузия почек, осложняющаяӯсяолигурией и почечной недостаточностью, яичники резко увеличены, отмечают ихразӯрывы и перекрут.Суммируя представленные данные, можно сказать, что, по-видимому, используемыев программах вспомогательӯной репродукции способы гормональной стимуляциифолликулогенеза и овуляции вносят свой вклад в увеӯличение числа такихошибок. Гибель зародышей в процессе имплантации может быть обусловленапереӯчисленными выше причинами, недостаточностью втоӯрой фазы цикла(изначально существовавшей или возӯникшей в процессе овариальной стимуляции),а такӯже, возможно, особенностями становления иммунологических отношенийэмбриона с организмом матери, вследствие отсутствия трубного периодаразвития, в который эти отношения начинают активно устанавлиӯваться («факторранней беременности» и т.п.).Трудно оценить влияние на гаметогенез и оплодотвоӯрение чисто внешних(экологических, антропогенных, социокультурных и т. п.) факторов. Однако они,безусӯловно, также вносят свой вклад в патологию раннего развития.Таким образом, в большинстве случаев пренатальные потери и приводящая к нимпатология у эмбриоӯнов при использовании методов вспомогательной реӯпродукциивозникают не необходимо и не случайно, а под влиянием определенных факторов,в том числе и приводящих к нарушению регулирующих репродукӯтивный процессмеханизмов. Элиминация же аноӯмальных гамет, зигот и эмбрионов - процесс какнеӯобходимый, так и закономерный, направленный на сохранение постоянствагенотипа популяции. ЗАКЛЮЧЕНИЕ Проблема бесплодия возникла отнюдь не сегодня, она сопровождает человечествоеще с древних времен. Современная медицина достаточно глубоко изучила причинывозникновения женского и мужского бесӯплодия. Разработаны как лекарственные,т.е. консерӯвативные, методы терапии бесплодия, так и оперативӯные.Однако неудовлетворенность достигнутыми резульӯтатами привела к разработкенового метода лечения бесплодия - ЭКО и ПЭ, который быстро завоевал поӯзициюлидирующего в этом направлении.В данной работе представлены основные проблемы программы ЭКО и ПЭ и возможныеосложнения, возникающие на различных ее этапах.Хотя применение метода ЭКО и ПЭ не позволяет в целом решить возникшую встране критическую деӯмографическую ситуацию, тем не менее широкое внедрениеего в практику здравоохранения поможет избавиться от бесплодия сотням тысячсупружеских пар, а следовательно, осуществить также их психолоӯгическуюреабилитацию.В заключение необходимо отметить, что благодаря успехам программы ЭКОдостигнуты положительные реӯзультаты в фундаментальных исследованияхчеловеӯческих гамет и эмбрионов. ЛИТЕРАТУРА 1. Экстракорпоральное оплодотворение и его новые направления в леченииженского бесплодия (теоретические и практические подходы): Руководство дляврачей / Под. ред. В.И.Кулакова, Б.В.Леонова – М., 2001, 782 с.2. Никитин А.И., Китаев Э.М., Савицкий Г.А., Иванова Р.Д., КалашниковаЕ.П. и Устинкина Т.И. Экстракорпоральное оплодотворение у человека споследующей имплантацией эмбриона и рождением ребенка. Арх. анатомии,гистологии и эмбриологии. – Л., 1987, Т.93, вып.10, с.39-43.3. Аншина М.Б., Здановский В.М. Если вам нужен ребенок. – М., 1998, 32с.4. Пшеничникова Т.Я. Бесплодие в браке. – М., 1991, 320с.5. Сметник В.П.. Тумилович Л.Г. Неоперативная гинекология. Руководстводля врачей. – М. 1999, с.476-480.
Дата добавления: 2015-08-08; просмотров: 1052;