Экология растений
Экология растений – наука об отношениях между растениями и окружающей средой. Среда, в которой живет растение, неоднородна и состоит из совокупности отдельных элементов, или факторов, значение которых для растений неодинаково. С этой точки зрения элементы среды делятся на три группы: 1) необходимые для существования растений; 2) вредные; 3) индифферентные (безразличные), не играющие никакой роли в жизни растений. Необходимые и вредные элементы среды в совокупности составляют экологические факторы. Индифферентные элементы к экологическим факторам не относятся.
Экологические факторы классифицируются по характеру воздействия на организм и по их происхождению. По характеру воздействия различают прямодействующие и косвеннодействующие экологические факторы. Прямодействующие факторы оказывают непосредственное влияние на растительный организм. Среди них особенно важную роль играют физиологически действующие факторы, например свет, вода, элементы минерального питания. Косвеннодействующими называют факторы, влияющие на организм опосредованно, через изменение прямодействующих факторов, например рельеф.
По происхождению выделяют следующие основные категории экологических факторов:
1. Абиотические факторы - факторы неживой природы:
а) климатические - свет, тепло, влага, состав и движение воздуха;
б) эдафические (почвенно-грунтовые) - разнообразные химические и физические свойства почв;
в) топографические (орографические) - факторы, обусловленные рельефом.
2. Биотические факторы - влияние друг на друга совместно обитающих организмов:
а) влияние на растения других (соседних) растений;
б) влияние на растения животных;
в) влияние на растения микроорганизмов.
3. Антропические (антропогенные) факторы - всевозможные воздействия на растения человека.
Экологические факторы влияют на растительный организм не изолированно друг от друга, а во всей их совокупности, образуя единую среду обитания. Различают две категории среды обитания - экотоп и местообитание (биотоп). Под экотопом понимают первичный комплекс абиотических экологических факторов на любом конкретном однородном участке земной поверхности. В чистом виде экотопы могут сформироваться лишь на участках, еще не заселенных организмами, например на недавно застывших лавовых потоках, на свежих осыпях обрывистых склонов, на речных песчаных и галечниковых отмелях. Под влиянием организмов, заселяющих экотоп, последний превращается в местообитание (биотоп), который представляет собой совокупность всех экологических факторов (абиотических, биотических, а нередко и антропических) на любом конкретном однородном участке земной поверхности.
Влияние экологических факторов на растительный организм очень разнообразно. Одни и те же факторы имеют неодинаковое значение для разных видов растений и на разных стадиях развития растений одного и того же вида.
Экологические факторы в природе соединены в комплексы, и на растение всегда действует весь комплекс факторов местообитания, причем общее влияние факторов местообитания на растение не равно сумме влияний отдельных факторов. Взаимодействие факторов проявляется в их частичной замещаемости, суть которой состоит в том, что уменьшение значений одного фактора может быть компенсировано увеличением интенсивности другого фактора, и поэтому реакция растения остается неизменной. В то же время ни один из необходимых растению экологических факторов не может быть полностью заменен другим: нельзя вырастить зеленое растение в полной темноте даже на очень плодородной почве или на дистиллированной воде при оптимальных условиях освещения.
Факторы, значения которых лежат вне зоны оптимума для данного вида, носят название лимитирующих. Именно лимитирующие факторы определяют существование вида в конкретном местообитании.
В отличие от животных растения ведут прикрепленный образ жизни и связаны в течение всей жизни с одними и теми же местообитаниями, которые подвергаются разнообразным изменениям во времени. Чтобы выжить, каждое растение должно обладать свойством приспособленности к определенному диапазону экологических условий, которое закрепляется наследственно и называется экологической пластичностью, или нормой реакции. Действие экологического фактора на растение можно изобразить графически в виде так называемой кривой жизнедеятельности, или экологической кривой (рис. 15.1).

Рис. 15.1. Схема действия экологического фактора на растение: 1 – точка минимума; 2 – точка оптимума; 3 – точка максимума.
На кривой жизнедеятельности выделяются три кардинальные точки: точка минимума и точка максимума, соответствующие крайним значениям фактора, при которых возможна жизнедеятельность организма; точка оптимума соответствует наиболее благоприятному значению фактора. Кроме того, на кривой жизнедеятельности выделяется несколько зон: зона оптимума - ограничивает диапазон благоприятных (комфортных) значений фактора; зоны пессимума - охватывают диапазоны резкого избытка и недостатка фактора, в пределах которых растение находится в состоянии сильного угнетения; зона жизнедеятельности располагается между экстремальными точками (минимумом и максимумом) и охватывает весь диапазон пластичности организма, в пределах которого организм способен отправлять свои жизненные функции и пребывать в активном состоянии. Вблизи экстремальных точек лежат сублетальные (крайне неблагоприятные) величины фактора, а за пределами – летальные (губительные).
Норма реакции определяется генотипом, чем больше протяженность кривой жизнедеятельности по оси абсцисс, тем выше экологическая пластичность растения или вида в целом.
Пластичность видов растений варьирует в широких пределах, в зависимости от этого они делятся на три группы: 1) стенотопы; 2) эвритопы; 3) умеренно пластичные виды. Стенотопами называют малопластичные виды, способные существовать в узком диапазоне того или иного экологического фактора, например растения влажных экваториальных лесов, которые обитают в условиях относительно стабильных температур, приблизительно от 20° до 30°С. Эвритопы характеризуются значительной пластичностью и способны осваивать разнообразные местообитания по отдельным факторам. К эвритопам относится, например, сосна обыкновенная (Pinus sylvestris), произрастающая на различных по увлажнению и плодородию почвах. Умеренно пластичные, к которым относится подавляющее большинство видов, занимают промежуточное положение между стенотопами и эвритопами. Разделяя виды на указанные выше группы, нужно учитывать, что эти группы выделяются по отдельным экологическим факторам и не характеризуют специфичности вида по остальным факторам. Вид может быть стенотопом по одному фактору, эвритопом по другому фактору и умеренно пластичным по отношению к третьему фактору.
Основной экологической единицей растительного мира является вид. Каждый вид объединяет особи, сходные по экологическим потребностям, и способен существовать лишь в определенных экологических условиях. Кривые жизнедеятельности разных видов могут налагаться в той или иной мере друг на друга, но полностью они никогда не совпадают. Это свидетельствует о том, что каждый вид растений экологически индивидуален и неповторим.
Однако вид - не единственная экологическая единица. В экологии растений широко используются такие категории, как экологическая группа и жизненная форма.
Экологическая группа отражает отношение растений к какому-либо одному фактору. Экологическая группа объединяет виды, одинаково реагирующие на тот или иной фактор, нуждающиеся для их нормального развития в сходных интенсивностях данного фактора и имеющие близкие значения точек оптимума. Виды, входящие в одну и ту же экологическую группу, характеризуются не только сходными потребностями в каком-то экологическом факторе, но также и рядом сходных наследственно закрепленных анатомо-морфологических признаков, обусловленных данным фактором. Наиболее важными экологическими факторами, влияющими на структуру растений, являются влажность и свет, большое значение имеют также температурный режим, особенности почв, конкурентные отношения в сообществе и ряд других условий. К сходным условиям растения могут приспосабливаться по-разному, вырабатывая разную “стратегию” использования имеющихся и компенсации недостающих жизненных факторов. Поэтому в пределах многих экологических групп можно найти растения, резко отличающиеся друг от друга по внешнему облику - габитусу и по анатомической структуре органов. Они имеют разную жизненную форму. Жизненная форма, в отличие от экологической группы, отражает приспособленность растений не к одному, отдельно взятому экологическому фактору, а ко всему комплексу условий местообитания.
Таким образом, в одну экологическую группу входят виды разных жизненных форм, и, наоборот, одна жизненная форма бывает представлена видами из разных экологических групп.
Экологические группы растений по отношению к влаге . Вода имеет исключительно важное значение для жизнедеятельности растительного организма. Протопласт живых клеток активен только в насыщенном водой состоянии, если он теряет определенное количество воды, клетка погибает. Передвижение веществ внутри растения осуществляется в виде водных растворов.
По отношению к влажности различают следующие основные группы растений.
1. Ксерофиты - растения, приспособившиеся к значительному постоянному или временному недостатку влаги в почве или в воздухе.
2. Мезофиты - растения, живущие в условиях достаточно умеренного увлажнения.
3. Гигрофиты - растения, обитающие при повышенной влажности атмосферы.
4. Гидрофиты - растения, приспособившиеся к водному образу жизни. В узком смысле гидрофитами называют только полупогруженные в воду растения, имеющие подводную и надводную части, или плавающие, т. е. живущие и в водной, и в воздушной среде. Полностью погруженные в воду растения называют гидатофитами.
Рассматривая типичные “средние” черты структуры листьев, стеблей и корней, мы, как правило, имеем в виду органы мезофитов, которые служат как бы эталоном.
Приспособление к более крайним условиям - недостатку или избытку влаги - вызывает те или иные уклонения от средней нормы.
Примерами гидатофитов могут служить элодея (Elodea), валлиснерия (Vallisneria), многие рдесты (Potamogeton), водяные лютики (Batrachium), уруть (Myriophyllum), роголистник (Ceratophyllum). Одни из них укореняются в грунте водоема, другие свободно взвешены в толще воды, и только во время цветения их соцветия выдвигаются над водой.
Строение гидатофитов определяется условиями жизни. Эти растения испытывают сильное затруднение с газообменом, так как в воде очень мало растворенного кислорода, причем его тем меньше, чем выше температура воды. Поэтому для гидатофитов характерна большая поверхность их органов по сравнению с общей массой. Листья у них тонкие, например у элодеи составлены всего двумя слоями клеток (рис. 15.2, А), и часто рассечены на нитевидные доли. Ботаники дали им меткое название - “листья-жабры”, что подчеркивает глубокое сходство рассеченных листьев с жаберными лепестками рыб, приспособленными к газообмену в водной среде.
К погруженным в воду растениям доходит ослабленный свет, так как часть лучей поглощается или отражается водой, и поэтому гидатофиты обладают некоторыми свойствами тенелюбов. В частности, в эпидерме присутствуют нормальные, фотосинтезирующие хлоропласты (рис. 15.2).
На поверхности эпидермы отсутствует кутикула, или она настолько тонка, что не представляет препятствия для прохождения воды, поэтому водные растения, вынутые из воды, полностью теряют воду и высыхают через несколько минут.
Вода гораздо плотнее воздуха и поэтому поддерживает погруженные в нее растения. К этому надо прибавить, что в тканях водных растений имеется много крупных межклетников, заполненных газами и образующих хорошо выраженную аэренхиму (рис. 15.2). Поэтому водные растения свободно взвешены в толще воды и не нуждаются в наличии особых механических тканей. Плохо развиты или совсем отсутствуют сосуды, так как растения воспринимают воду всей поверхностью тела.

Рис. 15.2. Анатомические особенности гидрофитов (поперечные срезы органов ): А – листовая пластинка гидатофита элодеи канадской (Elodea canadensis) сбоку от средней жилки; Б – сегмент листа гидатофита урути колосистой (Myriophyllum spicatum); В – пластинка плавающего листа аэрогидатофита кувшинки чисто-белой (Nymphaea candida); Г – стебель элодеи канадской (Elodea canadensis); Д – листовая пластинка гидатофита зостеры морской (Zostera marina); 1 – астросклереида; 2 – воздушная полость; 3 – гидатода; 4 – губчатый мезофилл; 5 – ксилема; 6 – паренхима первичной коры; 7 – мезофилл; 8 – проводящий пучок; 9 – палисадный мезофилл; 10 – склеренхимные волокна; 11 – устьице; 12 – флоэма; 13 - эпидерма.
Межклетники не только увеличивают плавучесть, но и способствуют регуляции газообмена. Днем, в процессе фотосинтеза, они заполняются кислородом, который в темное время суток используется для дыхания тканей; выделяющийся в процессе дыхания углекислый газ накапливается ночью в межклетниках, а днем используется в процессе фотосинтеза.
У большинства гидатофитов сильно развито вегетативное размножение, которое возмещает ослабленное семенное размножение.
Аэрогидатофиты - переходная группа. Ее составляют гидатофиты, у которых часть листьев плавает на поверхности воды, например кувшинка (Nymphaea), кубышка (Nuphar), водокрас (Hydrocharis), ряска (Lemna). Строение плавающих листьев отличается некоторыми особенностями (рис. 15.2, В ). Все устьица находятся на верхней стороне листа, т. е. направлены к атмосфере. Их очень много – у кубышки желтой (Nuphar lutea) на 1 мм2 поверхности их приходится до 650. Сильно развит палисадный мезофилл. Через устьица и по обширным межклетникам, развитым в пластинке листа и черешке, кислород поступает в корневища и корни, погруженные в грунт водоема.
Гидрофиты (аэрогидрофиты, «земноводные» растения) обычны по берегам водоемов, например аир болотный (Acorus calamus), стрелолист (Sagittaria), частуха (Alisma), камыш (Scirpus), тростник обыкновенный (Phragmites australis), хвощ приречный (Equisetum fluviatile), многие осоки (Carex) и др. В грунте водоема они образуют корневища с многочисленными придаточными корнями, а над поверхностью воды поднимаются или одни листья, или облиственные побеги.
Во всех органах гидрофитов имеется система хорошо развитых межклетных пространств, через которые снабжаются кислородом органы, погруженные в воду и в грунт водоема. Для многих гидрофитов характерна способность образовывать листья различной структуры в зависимости от того, в каких условиях протекает их развитие. Примером может служить стрелолист (рис. 15.3 ). Его лист, возвышающийся над водой, имеет прочный черешок и плотную стреловидную пластинку с хорошо выраженным палисадным мезофиллом; как в пластинке, так и в черешке находится система воздухоносных полостей.
Листья, погруженные в воду, имеют вид длинных и нежных лент без дифференциации на пластинку и черешок. Их внутреннее строение сходно со строением листьев типичных гидатофитов. Наконец, у одного и того же растения можно найти листья промежуточного характера с дифференцированной овальной пластинкой, плавающей на поверхности воды.
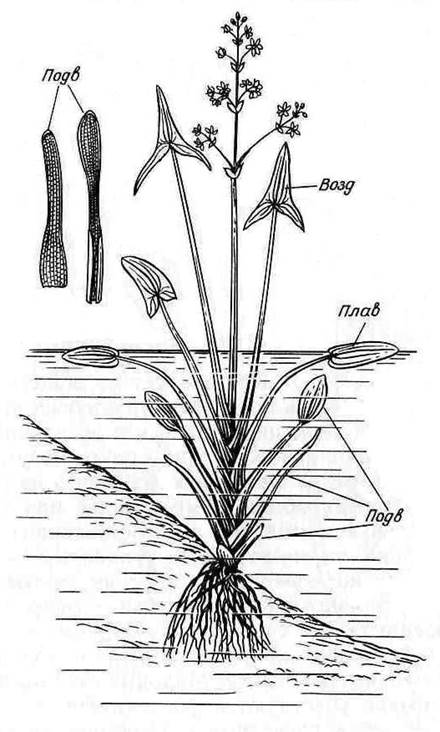
Рис. 15.3. Гетерофиллия у стрелолиста (Sagittaria sagittifolia): Подв - подводные; Плав – плавающие; Возд – воздушные листья.
В группу гигрофитов включают растения, обитающие на влажной почве, например на болотистых лугах или в сырых лесах. Поскольку эти растения не испытывают недостатка в воде, то в их структуре отсутствуют какие-либо особые приспособления, направленные на снижение транспирации. В листе медуницы (Pulmonaria) (рис. 15.4 ) клетки эпидермы тонкостенные, покрыты тонкой кутикулой. Устьица или находятся вровень с поверхностью листа, или даже приподняты над ней. Обширные межклетники создают общую большую испаряющую поверхность. Этому же способствует наличие рассеянных тонкостенных живых волосков. В условиях влажной атмосферы усиление транспирации ведет к лучшему передвижению растворов к побегам.
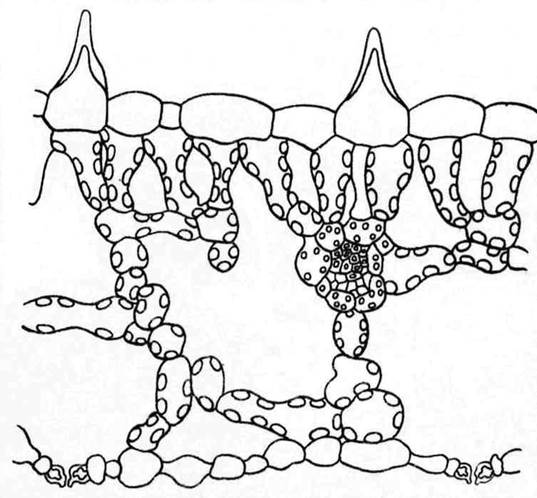
Рис. 15.4. Поперечный срез листа медуницы (Pulmonaria obscura).
У лесных гигрофитов к перечисленным признакам присоединяются черты, характерные для тенелюбивых растений.
Растения экологической группы ксерофитов в большинстве случаев имеют разнообразные приспособления к поддержанию водного баланса при недостатке почвенной и атмосферной влаги. В зависимости от основных путей приспособления к сухости местообитаний группа ксерофитов подразделяется на два типа: настоящие ксерофиты и ложные ксерофиты.
К настоящим ксерофитам относят такие растения, которые, произрастая на сухих местообитаниях, действительно испытывают недостаток влаги. Они обладают анатомо-морфологическими и физиологическими адаптациями. Совокупность всех анатомо-морфологических приспособлений настоящих ксерофитов придает им особую, так называемую ксероморфную структуру, которая отражает приспособление к уменьшению транспирации.
Ксероморфные признаки отчетливо проявляются в особенностях строения эпидермы. Основные клетки эпидермы у ксерофитов обладают утолщенными наружными стенками. Мощная кутикула покрывает эпидерму и заходит глубоко в устьичные щели (рис. 15.5). На поверхности эпидермы образуются восковые выделения в виде различных зерен, чешуек и палочек. На побегах восковой пальмы (Ceroxylon) толщина восковых выделений достигает5 мм.

Рис. 15.5. Поперечный срез листа алоэ (Aloe variegata) с погруженным устьицем.
К этим особенностям прибавляются различные виды трихомов. Густой покров из кроющих волосков снижает транспирацию непосредственно (замедляя движение воздуха на поверхности органов) и косвенно (отражая солнечные лучи и, тем самым, снижая нагревание побегов).
Для ксерофитов характерно погружение устьиц в ямки, так называемые крипты, в которых создается затишное пространство. Кроме того, стенки крипт могут иметь сложную конфигурацию. Например, у алоэ (рис. 15.5 ) выросты стенок клеток, почти смыкаясь между собой, создают дополнительное препятствие для выхода водяного пара из листа в атмосферу. У олеандра (Nerium oleander) в каждую большую крипту погружена целая группа устьиц, а полость крипты заполнена волосками, как бы заткнута ватной пробкой (рис. 15.6 ).
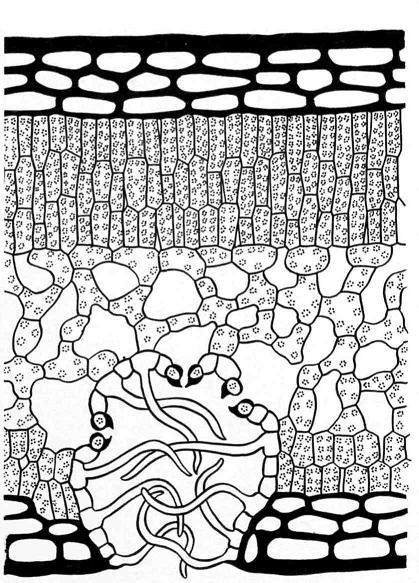
Рис. 15.6. Поперечный срез листа олеандра (Nerium oleander).
Внутренние ткани листьев у ксерофитов нередко отличаются мелкоклеточностью и сильной склерификацией, что приводит к сокращению межклетных пространств и общей внутренней испаряющей поверхности.
Ксерофиты, имеющие высокую степень склерификации, получили название склерофитов. Общая склерификация тканей часто сопровождается образованием по краю листа твердых шипов. Крайнее звено этого процесса - превращение листа или всего побега в твердую колючку.
Листья многих злаков имеют различные приспособления к свертыванию при недостатке влаги. У щучки (Deschampsia caespitosa) на нижней стороне листа, под эпидермой, залегает склеренхима, а все устьица находятся на верхней стороне листа. Они расположены на боковых сторонах гребней, идущих вдоль листовой пластинки. В углублениях, проходящих между гребнями, находятся моторные клетки - большие тонкостенные живые клетки, способные изменять объем. Если лист содержит достаточно воды, то моторные клетки, увеличивая объем, раскрывают лист. При недостатке воды моторные клетки уменьшаются в объеме, лист, как пружина, свертывается в трубку, и устьица оказываются внутри замкнутой полости (рис. 15.7 ).

Рис. 15.7. Поперечный срез листа щучки(Deschampsia caespitosa): 1 – часть пластинки листа при большом увеличении; 2 – срез всей пластинки листа; 3 – пластинка листа в свернутом состоянии; МК – моторные клетки; ПП - проводящий пучок; Скл – слеренхима; Хл – хлоренхима; Э – эпидерма.
Редукция листьев характерна для многих кустарников Средиземноморья, пустынь Средней Азии и других мест с сухим и жарким летом: джузгуна (Calligonum), саксаула (Haloxylon), испанского дрока (Spartium), эфедры (Ephedra) и многих других. У этих растений стебли принимают на себя функцию фотосинтеза, а листья или недоразвиваются, или опадают рано весной. В стеблях под эпидермой находится хорошо развитая палисадная ткань (рис. 15.8).
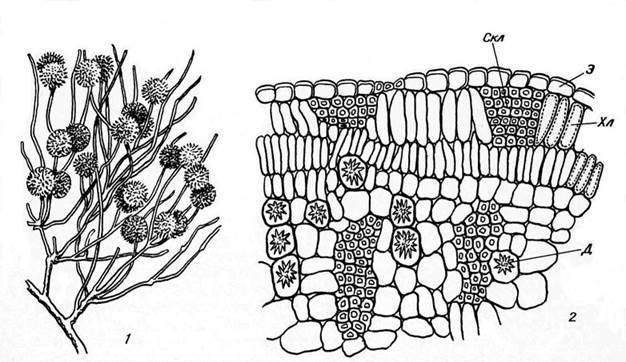
Рис. 15.8. Ветка джузгуна (Calligonum) (1) и часть ее поперечного среза (2): Д – друза; Скл – склеренхима; Хл – хлоренхима; Э – эпидерма.
Поскольку ксерофиты большей частью растут в степях, пустынях, на сухих склонах и других открытых местах, они в равной мере приспособлены к яркому освещению. Поэтому не всегда можно разграничить ксероморфные признаки и признаки, вызванные приспособлением к яркому освещению.
Однако главными адаптациями настоящих ксерофитов к сухости местообитаний являются физиологические особенности: высокое осмотическое давление клеточного сока и засухоустойчивость протопласта.
К ложным ксерофитам относятся растения, которые произрастают на сухих местообитаниях, но не испытывают недостатка влаги. Ложные ксерофиты имеют приспособления, позволяющие добывать достаточное количество воды, и, образно говоря, “убегать от засухи”. Поэтому у них ослабляются или отсутствуют совсем признаки ксероморфной структуры.
К группе ложных ксерофитов, прежде всего, относятся пустынно-степные суккуленты. Суккулентами называют сочные мясистые растения с сильно развитой водоносной тканью в надземных или подземных органах. Различают две основные жизненные формы - стеблевые и листовые суккуленты. Стеблевые суккуленты имеют толстый, сочный стебель различной формы. Листья всегда редуцированы и превращены в колючки. Типичными представителями стеблевых суккулентов являются кактусы и кактусовидные молочаи. У листовых суккулентов водоносная ткань развивается в листьях, которые становятся толстыми и сочными, и в которых накапливается много воды. Стебель у них сухой и жесткий. Типичными листовыми суккулентами являются виды алоэ (Aloe) и агавы (Agave).
В благоприятные периоды, когда почва увлажняется выпавшими атмосферными осадками, суккуленты, обладающие сильно разветвленной поверхностной корневой системой, быстро накапливают в своих водоносных тканях большое количество воды и затем во время последующей длительной засухи очень экономно расходуют ее, практически не испытывая недостатка влаги. Экономия воды осуществляется благодаря целому ряду адаптивных признаков: устьица у суккулентов малочисленны, располагаются в углублениях и открываются только ночью, когда температура понижается, а влажность воздуха повышается; клетки эпидермы покрыты толстой кутикулой и восковым налетом. Все это обусловливает очень низкую интенсивность общей транспирации у суккулентов и позволяет им осваивать крайне сухие местообитания.
Однако свойственный суккулентам тип водного обмена затрудняет газообмен и поэтому не обеспечивает достаточной интенсивности фотосинтеза. Устьица у этих растений открыты только ночью, когда процесс фотосинтеза невозможен. Углекислый газ запасается ночью в вакуолях, связываясь в виде органических кислот, а затем днем отщепляется и используется в процессе фотосинтеза. В связи с этим интенсивность фотосинтеза у суккулентов очень низка, накопление биомассы и рост у них протекают медленно, что обусловливает низкую конкурентную способность этих растений.
К ложным ксерофитам относятся также пустынно-степные эфемеры и эфемероиды. Это растения с очень коротким периодом вегетации, приуроченным к более прохладному и влажному сезону года. За этот короткий (иногда не более 4-6 недель) благоприятный срок они успевают пройти весь годичный цикл развития (от прорастания до образования семян), а остальную неблагоприятную часть года переживают в состоянии покоя. Такая ритмика сезонного развития позволяет эфемерам и эфемероидам “убегать от засухи во времени”.
К эфемерам относят однолетние растения, которые неблагоприятный период переживают в виде семян и размножаются только семенами. Обычно они имеют малые размеры, так как за короткий срок не успевают сформировать значительную вегетативную массу. Эфемероиды являются многолетними растениями. Поэтому они переживают неблагоприятное время не только в виде семян, но и в виде покоящихся подземных органов - луковиц, корневищ, клубней.
Поскольку эфемеры и эфемероиды приурочивают свой активный период к влажному сезону года, они не испытывают дефицита влаги. Поэтому для них характерна, как и для мезофитов, мезоморфная структура. Однако их семена и подземные органы характеризуются высокой засухоустойчивостью и жаростойкостью.
Глубококорневые ложные ксерофиты “убегают от засухи в пространстве”. Эти растения имеют очень глубокие корневые системы (до 15-20 м и более), которые проникают к водоносным горизонтам грунта, где интенсивно разветвляются и бесперебойно снабжают растение водой даже в периоды самой сильной засухи. Не испытывая обезвоживания, глубококорневые ложные ксерофиты сохраняют в целом мезоморфный облик, хотя у них наблюдается некоторое уменьшение общей испаряющей поверхности за счет превращения части листьев или побегов в колючки. Типичным представителем данной жизненной формы является верблюжья колючка (Alhagi pseudalhagi) из семейства бобовых, которая образует заросли в пустынях Средней Азии и Казахстана.
Экологические группы растений по отношению к свету . Свет имеет в жизни растений очень большое значение. Прежде всего, он является необходимым условием фотосинтеза, в процессе которого растения связывают световую энергию и за счет этой энергии осуществляют синтез органических веществ из углекислого газа и воды. Свет оказывает также влияние на ряд других жизненных функций растений: прорастание семян, рост, развитие репродуктивных органов, транспирацию и др. Кроме того, с изменением условий освещения изменяются некоторые другие факторы, например температура воздуха и почвы, их влажность, и, таким образом, свет оказывает на растения не только прямое, но и косвенное воздействие.
Количество и качество света в местообитаниях изменяются в зависимости от географических факторов (географической широты и высоты над уровнем моря), а также под влиянием местных факторов (рельефа и затенения, создаваемого совместно произрастающими растениями). Поэтому в процессе эволюции образовались виды растений, нуждающиеся в разных условиях освещения. Обычно выделяют три экологические группы растений: 1) гелиофиты – светолюбивые растения; 2) сциогелиофиты - теневыносливые растения; 3) сциофиты - тенелюбивые растения.
Гелиофиты, или светолюбивые растения, - это растения открытых (незатененных) местообитаний. Они встречаются во всех природных зонах Земли. Гелиофитами являются, например, многие виды растений верхних ярусов степей, лугов и лесов, наскальные мхи и лишайники, многие виды разреженной пустынной, тундровой и высокогорной растительности.
Побеги светолюбивых растений довольно толстые, с хорошо развитой ксилемой и механической тканью. Междоузлия укорочены, типично значительное ветвление, в результате чего нередко возникает розеточность и образование формы роста типа “подушка”.
Листья гелиофитов в целом имеют более мелкие размеры и располагаются в пространстве так, что в самые яркие полуденные часы солнечные лучи как бы “скользят” по листовой пластинке и меньше усваиваются, а в утренние и вечерние часы падают на ее плоскость, используясь максимально.
Анатомические особенности строения листа у гелиофитов также направлены на уменьшение поглощения света. Так, листовые пластинки многих светолюбивых растений имеют специфическую поверхность: либо блестящую, либо покрытую восковым налетом, либо густо опушенную светлыми волосками. Во всех этих случаях листовые пластинки способны отражать значительную часть солнечного света. Кроме того, у гелиофитов хорошо развиты эпидерма и кутикула, которые сильно затрудняют проникновение света в мезофилл листа. Установлено, что эпидерма светолюбивых растений пропускает не более 15% падающего света.
Мезофилл листа имеет плотное строение за счет сильного развития палисадной паренхимы, образующейся как у верхней, так и у нижней стороны листа (рис. 15.6 ).
Хлоропласты у гелиофитов мелкие, они густо наполняют клетку, частично затеняя друг друга. В составе хлорофилла преобладает более светоустойчивая форма “а” над формой “в” (а/в = 4.5-5.5). Общее содержание хлорофилла невысокое - 1.5-3 мг на 1 г сухой навески листа. Поэтому листья гелиофитов обычно имеют светло-зеленую окраску.
Сциогелиофитами называют теневыносливые растения, которые обладают высокой пластичностью по отношению к свету и могут нормально развиваться как при полном освещении, так и в условиях более или менее выраженного затенения. К теневыносливым растениям относятся большинство лесных растений, многие луговые травы и небольшое число степных, тундровых и некоторых других растений.
Сциофиты нормально растут и развиваются в условиях слабого освещения, отрицательно реагируя на прямой солнечный свет. Поэтому их по праву можно назвать тенелюбивыми растениями. К этой экологической группе относятся растения нижних ярусов густых тенистых лесов и густотравных лугов, погруженные в воду растения, немногочисленные обитатели пещер.
Приспособления тенелюбов к свету во многом противоположны адаптациям светолюбивых растений. Листья сциофитов в целом более крупные и более тонкие, чем у гелиофитов, в пространстве они ориентируются так, чтобы получить максимум света. Для них характерно отсутствие или слабое развитие кутикулы, отсутствие опушения и воскового налета. Поэтому свет проникает в лист сравнительно легко - эпидерма тенелюбов пропускает до 98% падающего света. Мезофилл рыхлый, крупноклеточный, не дифференцирован (или слабо дифференцирован) на столбчатую и губчатую паренхиму (рис. 15.4 ).
Хлоропласты у тенелюбов крупные, но их мало в клетке, и поэтому они не затеняют друг друга. Отношение содержания форм хлорофилла “а” и “в” снижается (а/в = 2.0-2.5). Общее содержание хлорофилла довольно велико - до 7-8 мг/1 г листа. Поэтому листья сциофитов имеют, как правило, темно-зеленую окраску.
У водных тенелюбов хорошо выражено адаптивное изменение состава фотосинтезирующих пигментов в зависимости от глубины обитания, а именно: у высших водных растений и у зеленых водорослей, обитающих в верхнем слое воды, преобладают хлорофиллы, у цианобактерий (синезеленых водорослей) к хлорофиллу прибавляется фикоцианин, у бурых водорослей - фукоксантин, у самых глубоководных красных водорослей - фикоэритрин.
Своеобразным типом физиологической адаптации некоторых тенелюбов к недостатку света является утрата способности к фотосинтезу и переход к гетеротрофному питанию. Это растения – симбиотрофы (микотрофы), получающие органические вещества с помощью грибов-симбионтов (подъельник (Hypopitys monotropa) из семейства вертляницевых, ладьян (Corallorhiza), гнездовка (Neottia), надбородник (Epipogium) из семейства орхидных). Побеги этих растений теряют зеленую окраску, листья редуцируются и превращаются в бесцветные чешуи. Корневая система приобретает своеобразную форму: под влиянием гриба рост корней в длину ограничивается, зато они разрастаются в толщину (рис. 15.9 ).

Рис. 15.9. Растения – микотрофы : 1 - корни ладьяна трехнадрезного (Corallorhiza trifida); 2 - гнездовка настоящая (Neottia nidus-avis); 3 - подъельник обыкновенный (Hypopitys monotropa).
Растения - паразиты других высших растений многочисленны в нижних ярусах густых влажнотропических лесов. Классическим примером является раффлезия Арнольда (Rafflesia arnoldii), которая не имеет ни корней, ни листьев. Внутри растения-хозяина развиваются присоски-гаустории, а снаружи формируется огромный цветок, достигающий в диаметре 1 м и весящий 4- 6 кг. Встречаются паразиты и среди растений умеренных широт (виды заразихи (Orobanche), петров крест (Lathraea squamaria), повилика (Cuscuta)). Они полностью утрачивают зеленую окраску; стебли и листья их крайне редуцированы; вместо корней формируются присоски-гаустории, внедряющиеся в ткани растения-хозяина и соединяющиеся с его проводящей системой.
Кроме настоящих паразитов, полностью утративших способность к фотосинтезу, встречаются разнообразные формы полупаразитов. Полупаразиты сохраняют нормальные зеленые облиственные побеги, а от растения-хозяина получают, главным образом, воду и минеральные вещества: у них вместо мелких сосущих корней образуются присоски, внедряющиеся в ткани корней или ветвей растения-хозяина. К полупаразитам принадлежит широко распространенный в Западной Европе кустарничек омела (Viscum album), поселяющаяся в кронах деревьев, а также многие луговые растения из семейства норичниковых (погремок (Rhinanthus), марьянник (Melampyrum), очанка (Euphrasia)) (рис. 15.10).
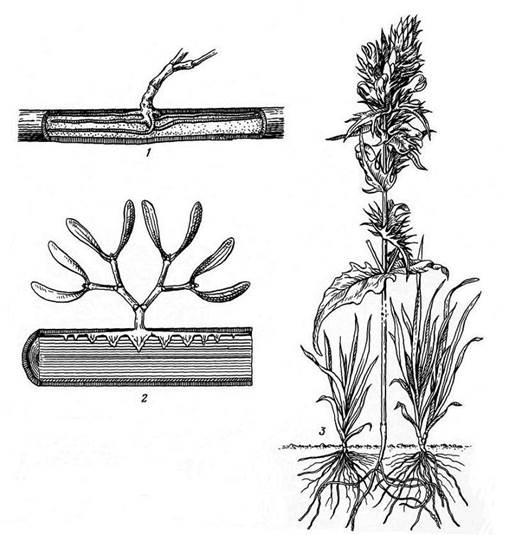
Рис. 15.10. Растения-полупаразиты : 1 – омела (Viscum album), нижняя часть растения; видны видоизмененные корни под корой растения-хозяина; 2 – продольный разрез ветви растения-хозяина; видны присоски на корнях омелы; 3 – марьянник полевой (Melampyrum arvense), паразитирующий на луговых злаках.
В условиях глубокого затенения нижних ярусов влажнотропических лесов выработались особые жизненные формы растений, выносящих в конечном счете основную массу побегов, вегетативных и цветоносных, в верхние ярусы, к свету. Это удается благодаря специфическим способам роста. Сюда относятся лианы и эпифиты.
Лианы выбираются на свет, используя как опору соседние растения, скалы и другие твердые предметы. Поэтому их еще называют лазящими растениями в широком смысле. Лианы могут быть древесными и травянистыми и наиболее характерны для влажнотропических лесов. В умеренной зоне их больше всего во влажных ольшаниках по берегам водоемов; это почти исключительно травы, такие как хмель (Humulus lupulus), калистегия (Calystegia), ясменник (Asperula) и т. д. В лесах Кавказа встречается довольно много деревянистых лиан (сассапариль (Smilax), обвойник (Periploca), ежевики). На Дальнем Востоке они представлены лимонником китайским (Schisandra chinensis), актинидией (Actinidia), виноградом (Vitis).
Специфика роста лиан состоит в том, что сначала их стебли растут очень быстро, а листья отстают и остаются несколько недоразвитыми. Когда же, использовав опору, растение выносит верхние побеги на свет, там развиваются нормальные зеленые листья и соцветия. Анатомическая структура стеблей лиан резко отличается от типичной структуры прямостоячих стеблей и отражает специфику стебля, наиболее гибкого даже при значительном одревеснении (у древесных лиан). В частности, стебли лиан обычно имеют пучковое строение и широкие паренхимные лучи между пучками.
Эпифиты - это особая жизненная форма растений, встречающаяся преимущественно во влажнотропических лесах. Чаще всего это травянистые растения, которые поселяются на стволах и ветвях других видов, но не являются паразитами по отношению к ним, а лишь “снимают квартиру” высоко над землей, в условиях более благоприятного освещения. Естественно, что это возможно лишь при очень высокой влажности воздуха. Воздушные корни многих эпифитов поглощают водяные пары с помощью специальной всасывающей ткани - веламена (см. раздел 4). Сначала маленькое растение, выросшее на чужой ветви из семени, практически может жить только за счет воздушного питания. Позднее около него накапливается некоторое количество опада, мусора, и создается собственная “почва”, чему способствуют у некоторых эпифитов специализированные листья, плотно прижатые к субстрату, например у папоротника олений рог (Platycerium).
Интересную жизненную форму представляют также эфемеры и эфемероиды листопадных лесов, например кандык сибирский (Erythronium sibiricum), прострел раскрытый (Pulsatilla patens), горицвет весенний (Adonis vernalis), ветреница лесная (Anemone sylvestris), медуница мягчайшая (Pulmonaria dacica). Все они являются светолюбивыми растениями и могут произрастать в нижних ярусах леса только благодаря тому, что сдвигают свой короткий вегетационный период на весну и начало лета, когда листва на деревьях еще не успевает распуститься, и освещенность у поверхности почвы оказывается высокой. Ко времени полного распускания листьев в кронах деревьев и появления затенения они успевают отцвести и образовать плоды.
Экологические группы по отношению к температуре . Тепло является одним из необходимых условий существования растений, так как все физиологические процессы и биохимические реакции зависят от температуры. Поэтому нормальный рост и развитие растений осуществляются лишь при наличии определенного количества тепла и определенной продолжительности его воздействия.
Выделяют четыре экологические группы растений: 1) мегатермы - жаростойкие растения; 2) мезотермы - теплолюбивые, но не жаростойкие растения; 3) микротермы - не требовательные к теплу растения, произрастающие в условиях умеренно холодного климата; 4) гекистотермы - особо холодостойкие растения. Последние две группы нередко объединяют в одну группу холодостойких растений.
Мегатермы имеют целый ряд анатомо-морфологических, биологических и физиологических приспособлений, которые позволяют им нормально отправлять свои жизненные функции при сравнительно высоких температурах.
К анатомо-морфологическим особенностям мегатермов относятся: а) густое белое или серебристое опушение или блестящая поверхность листьев, отражающие значительную часть солнечной радиации; б) уменьшение поверхности, поглощающей солнечную радиацию, которое достигается редукцией листьев, свертыванием листовых пластинок в трубку, поворачиванием листовых пластинок ребром к солнцу и другими способами; в) сильное развитие покровных тканей, изолирующих внутренние ткани растений от высоких температур окружающей среды. Эти особенности предохраняют жаростойкие растения от перегрева, имея в то же время адаптивное значение и против иссушения, которое обычно сопутствует высоким температурам.
Из биологических (поведенческих) приспособлений следует отметить явление так называемого “убегания” от экстремально высоких температур. Так, пустынные и степные эфемеры и эфемероиды значительно сокращают свой вегетационный период и приурочивают его к более прохладному сезону, тем самым “убегая во времени” не только от засухи, но и от высоких температур.
Особенно важное значение для жаростойких растений имеют физиологические адаптации, прежде всего способность протопласта без вреда переносить высокие температуры. Для некоторых растений характерна высокая интенсивность транспирации, ведущая к охлаждению тела и предохраняющая их от перегрева.
Жаростойкие растения характерны для сухих и жарких областей земного шара, также как и рассмотренные ранее ксерофиты. Кроме того, к мегатермам относятся наскальные мхи и лишайники освещенных местообитаний различных широт и виды бактерий, грибов и водорослей, обитающих в горячих источниках.
К типичным мезотермам относятся растения влажнотропического пояса, которые живут в условиях постоянно теплого, но не жаркого климата, в диапазоне температур 20-30°С. Как правило, эти растения не имеют никаких приспособлений к температурному режиму. К мезотермам умеренных широт относятся так называемые широколиственные древесные породы: бук (Fagus), граб (Carpinus), каштан (Castanea) и др., а также многочисленные травы из нижних ярусов широколиственных лесов. Эти растения тяготеют в своем географическом распространении к океаническим окраинам материков с мягким влажным климатом.
Микротермы - умеренно холодостойкие растения - характерны для бореально-лесной области, к наиболее холодостойким растениям - гекистотермам - относятся тундровые и высокогорные растения.
Основную адаптивную роль у холодостойких растений играют физиологические механизмы защиты: прежде всего, понижение точки замерзания клеточного сока и так называемая “льдоустойчивость”, под которой понимают способность растений без вреда переносить образование льда в их тканях, а также переход многолетних растений в состояние зимнего покоя. Именно в состоянии зимнего покоя растения обладают наибольшей холодостойкостью.
Для наиболее холодостойких растений - гекистотермов большое адаптивное значение имеют такие морфологические особенности, как небольшие размеры и специфичные формы роста. Действительно, подавляющее большинство тундровых и высокогорных растений имеют малые (карликовые) размеры, например береза карликовая (Betula nana), ива полярная (Salix polaris) и др. Экологическое значение карликовости заключается в том, что растение располагается в более благоприятных условиях, летом лучше прогревается солнцем, а зимой защищается снежным покровом. Исследователи арктических областей уже давно обратили внимание на то, что торчащие над снегом верхние части тундровых кустарников зимой в большинстве случаев отмерзают или истираются в порошок снежными, ледяными и минеральными частицами, которые переносятся частыми и сильными ветрами. Таким образом, все, что располагается выше поверхности снега, здесь обречено на гибель.
Подобное экологическое значение имеет возникновение таких своеобразных форм роста, как стланцы и растения-подушки. Стланцы - это стелющиеся формы деревьев, кустарников и кустарничков, например кедровый стланик (Pinus pumila), багульник стелющийся (Ledum decumbens), полярные виды водяники (Empetrum), можжевельник туркестанский (Juniperus turkestanica) и др.
Растения-подушки (см. раздел 4) образуются в результате сильного ветвления и крайне медленного роста надземных побегов. Между побегами накапливаются растительный опад и минеральные частицы. Все это ведет к образованию компактной и достаточно плотной формы роста. По некоторым растениям-подушкам можно ходить, как по твердой почве. Экологический смысл подушковидной формы роста состоит в следующем. Благодаря компактной структуре растения-подушки успешно противостоят холодным ветрам. Поверхность их нагревается почти так же, как поверхность почвы, а колебания температуры внутри подушки не так велики, как в окружающей среде. Поэтому внутри растения-подушки, как в парнике, сохраняется более благоприятный температурный и водный режим. Кроме того, непрерывное накопление в подушке растительного опада и его дальнейшее разложение способствуют повышению плодородия почвы под ней.
Подушковидные формы роста образуют в соответствующих условиях травянистые, полудревесные и древесные растения различных семейств: бобовых, розоцветных, зонтичных, гвоздичных, первоцветных и др. Подушки очень распространены и иногда целиком определяют ландшафт в высокогорьях всех материков, а также на каменистых океанических островах, особенно в Южном полушарии, на морских побережьях, в арктических тундрах и т.д. Некоторые подушки имеют резко выраженные внешние черты ксероморфизма, в частности колючки различного происхождения.
Экологические группы по отношению к почвенным факторам. Почва является одной из важнейших сред жизни сухопутных растений. Она служит субстратом для закрепления растений на определенном месте, а также представляет собой питательную среду, из которой растения усваивают воду и элементы минерального питания. Во всем многообразии почвенно-грунтовых факторов принято различать химические и физические свойства почвы. Из химических свойств почвенной среды основное экологическое значение имеют реакция почвенной среды и солевой режим почв.
В природных условиях реакция почвы складывается под влиянием климата, почвообразующей породы, грунтовых вод и растительности. Разные виды растений неодинаково реагируют на реакцию почвы и, с данной точки зрения, разделяются на три экологические группы: 1) ацидофиты; 2) базифиты и 3) нейтрофиты.
К ацидофитам относят растения, предпочитающие кислые почвы. Ацидофитами являются растения сфагновых болот, например сфагновые мхи (Sphagnum), багульник болотный (Ledum palustre), кассандра, или болотный мирт (Chamaedaphne calyculata), подбел (Andromeda polifolia), клюква (Oxycoccus); некоторые лесные и луговые виды, например брусника (Vaccinium vitis-idaea), черника (Vaccinium myrtillus), хвощ лесной (Equisetum sylvaticum).
К базифитамотносят растения, предпочитающие почвы, богатые основаниями и потому имеющие щелочную реакцию. Базифиты произрастают на карбонатных и солонцовых почвах, а также на обнажениях карбонатных пород.
Нейтрофиты предпочитают почвы с нейтральной реакцией. Однако многие нейтрофиты имеют широкие зоны оптимума - от слабокислой до слабощелочной реакции.
Под солевым режимом почв понимают состав и количественные соотношения химических веществ в почве, которые определяют содержание в ней элементов минерального питания. Растения реагируют на содержание как отдельных элементов минерального питания, так и всей их совокупности, которая определяет уровень плодородия почвы (или ее “трофность”). Разные виды растений нуждаются для своего нормального развития в различных количествах минеральных элементов в почве. В соответствии с этим различают три экологические группы: 1) олиготрофы; 2) мезотрофы; 3) эвтрофы (мегатрофы).
Олиготрофами называют растения, довольствующиеся очень малым содержанием элементов минерального питания. Типичными олиготрофами являются растения сфагновых болот: сфагновые мхи, багульник, подбел, клюква и др. Из древесных пород к олиготрофам относится сосна обыкновенная, а из луговых растений - белоус (Nardus stricta).
Мезотрофы - это растения, умеренно требовательные к содержанию элементов минерального питания. Они произрастают на небогатых, но и не на очень бедных почвах. К мезотрофам относятся многие древесные породы – кедр сибирский (Pinus sibirica), пихта сибирская (Abies sibirica), береза повислая (Betula pendula), осина (Populus tremula), многие таежные травы – кисличка (Oxalis acetosella), вороний глаз (Paris quadrifolia), седмичник (Trientalis europaea) и др.
Эвтрофы предъявляют высокие требования к содержанию элементов минерального питания, поэтому произрастают на высокоплодородных почвах. К эвтрофам относятся большинство степных и луговых растений, например ковыль перистый (Stipa pennata), тонконог (Koeleria cristata), пырей ползучий (Elytrigia repens), а также некоторые растения низинных болот, например тростник обыкновенный (Phragmites australis).
У представителей данных экологических групп не обнаруживается каких-либо специфических анатомо-морфологических приспособительных признаков, обусловленных трофностью их местообитаний. Однако олиготрофы часто обладают ксероморфными признаками, такими как мелкие жесткие листья, толстая кутикула и т. п. Очевидно, морфологическая и анатомическая реакция на недостаток почвенного питания сходна с некоторыми типами реакций на недостаток влаги, что вполне объяснимо с точки зрения ухудшения условий роста в том и другом случае.
Некоторые автотрофные растения, обитающие, как правило, на болотах (в тропической и отчасти в умеренной зоне), восполняют недостаток азота в субстрате дополнительным питанием за счет мелких животных, в частности насекомых, тела которых перевариваются при помощи ферментов, выделяемых специальными железками на листьях насекомоядных, или хищных, растений. Обычно способность к такому виду питания сопровождается формированием разнообразных ловчих приспособлений.
У обычной на сфагновых болотах росянки (Drosera rotundifolia, рис. 15.11, 1) листья покрыты красноватыми железистыми волосками, выделяющими на кончиках капельки липкого блестящего секрета. Мелкие насекомые прилипают к листу и своими движениями раздражают другие железистые волоски листа, которые медленно загибаются и плотно окружают насекомое своими железками. Растворение и всасывание пищи происходит в течение нескольких дней, после чего волоски расправляются, и лист снова может ловить добычу.
Ловчий аппарат венериной мухоловки (Dionaea muscipula), живущей на торфяниках востока Северной Америки, имеет сложное строение (рис. 15.11, 2, 3). На листьях имеются чувствительные щетинки, которые вызывают резкое захлопывание двух лопастей пластинки при прикосновении насекомого.
Ловчие листья у непентесов (Nepenthes, рис. 15.11, 4), лазящих растений прибрежных тропических зарослей Индо-Малайской области, имеют длинный черешок, нижняя часть которого широкая, пластинчатая, зеленая (фотосинтезирующая); средняя - суженная, стеблеподобная, вьющаяся (она и обвивает опору), а верхняя превращена в пестрый кувшинчик, прикрытый сверху крышечкой - листовой пластинкой. По краю кувшинчика выделяется сахаристая жидкость, привлекающая насекомых. Попав в кувшинчик, насекомое соскальзывает по гладкой внутренней стенке на его дно, где находится переваривающая жидкость.
В стоячих водоемах у нас обычно погруженное плавающее растение пузырчатка (Utricularia, рис. 15.11, 5, 6). Она не имеет корней; листья рассечены на узкие нитевидные дольки, на концах которых находятся ловчие пузырьки с клапаном, открывающимся внутрь. Мелкие насекомые или рачки не могут выбраться из пузырька и перевариваются там.
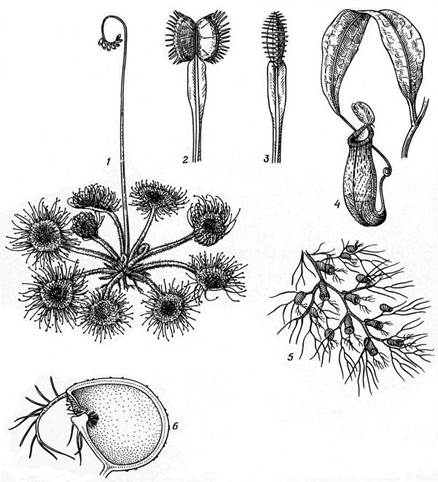
Рис. 15.11. Насекомоядные растения: 1 – росянка (Drosera rotundifolia); 2 и 3 – венерина мухоловка (Dionaea muscipula), открытый и закрывшийся лист; 4 – непентес (Nepenthes), лист-«кувшинчик»; 5 и 6 – пузырчатка (Utricularia), часть листа и ловчий пузырек.
Для большинства растений вредным является как недостаточное, так и избыточное содержание минеральных элементов. Однако некоторые растения приспособились к избыточно высокому содержанию элементов питания. Наиболее изученными являются следующие четыре группы.
1. Нитрофиты - растения, адаптированные к избыточному содержанию азота. Типичные нитрофиты произрастают на мусорных и навозных кучах и отвалах, на захламленных вырубках, заброшенных усадьбах и других местообитаниях, где идет усиленная нитрификация. Они поглощают нитраты в таких количествах, что их можно обнаружить даже в клеточном соке данных растений. К нитрофитам относятся крапива двудомная (Urtica dioica), яснотка белая (Lamium album), виды лопуха (Arctium), малина (Rubus idaeus), бузина (Sambucus) и др.
2. Кальцефиты - растения, приспособленные к избыточному содержанию в почве кальция. Они произрастают на карбонатных (известковых) почвах, а также на обнажениях известняков и мела. К кальцефитам относятся многие лесные и степные растения, например венерин башмачок (Cypripedium calceolus), ветреница лесная (Anemone sylvestris), люцерна серповидная (Medicago falcata) и т. д. Из древесных пород кальцефитами являются лиственница сибирская (Larix sibirica), бук (Fagus sylvatica), дуб пушистый (Quercus pubescens) и некоторые другие. Особенно разнообразен состав кальцефитов на известковых и меловых обнажениях, которые образуют особую, так называемую “меловую” флору.
3. Токсикофиты объединяют виды, устойчивые к высоким концентрациям определенных тяжелых металлов (Zn, Pb, Cr, Ni, Co, Cu) и способные даже накапливать ионы этих металлов. Токсикофиты приурочены в своем распространении к почвам, формирующимся на породах, богатых элементами тяжелых металлов, а также к отвалам пустой породы промышленных разработок залежей данных металлов. Типичными токсикофитами-концентраторами, пригодными для индикации почв, содержащих много свинца, являются овсяница овечья (Festuca ovina), полевица тонкая (Agrostis tenuis); на цинковых почвах - фиалка (Viola calaminaria), ярутка полевая (Thlaspi arvense), некоторые виды смолевки (Silene); на почвах, богатых селеном, - ряд видов астрагала (Astragalus); на почвах, богатых медью, - оберна (Oberna behen), качим (Gypsophila patrinii), виды шпажника (Gladiolus) и т. д.
4. Галофиты - растения, устойчивые к высокому содержанию ионов легкорастворимых солей. Избыток солей увеличивает концентрацию почвенного раствора, в результате чего создаются трудности в поглощении растениями питательных веществ. Галофиты поглощают эти вещества благодаря повышенному осмотическому давлению клеточного сока. К жизни на засоленных почвах разные галофиты приспособились по-разному: одни из них выделяют избыток поглощенных из почвы солей или через особые железки на поверхности листьев и стеблей (кермек (Limonium gmelinii), млечник (Glaux maritima)), или сбрасывая листья и веточки по мере накопления в них предельных концентраций солей (подорожник солончаковый (Plantago maritima), гребенщик (Tamarix)). Другие галофиты являются суккулентами, что способствует снижению концентрации солей в клеточном соке (солерос (Salicornia europaea), виды солянок (Salsola)). Главной особенностью галофитов является физиологическая устойчивость протопласта их клеток к ионам солей.
Из физических свойств почвы основное экологическое значение имеют воздушный, водный и температурный режимы, механический состав и структура почвы, ее пористость, твердость и пластичность. Воздушный, водный и температурный режимы почвы определяются климатическими факторами. Остальные физические свойства почвы оказывают на растения, в основном, косвенное влияние. И только на песчаных и очень твердых (каменистых) субстратах растения находятся под прямым воздействием некоторых их физических свойств. В результате формируются две экологические группы - псаммофиты и петрофиты (литофиты).
В группу псаммофитов объединяют растения, приспособленные к жизни на подвижных песках, которые лишь условно можно назвать почвами. Такого рода субстраты занимают огромные пространства в песчаных пустынях, а также встречаются по берегам морей, крупных рек и озер. Специфической экологической особенностью песков является их сыпучесть. В результате в жизни псаммофитов постоянно существует угроза либо засыпания песком надземных частей растений, либо, напротив, выдувания песка и оголения их корней. Именно этот экологический фактор обусловливает основные анатомо-морфологические и биологические адаптивные признаки, свойственные псаммофитам.
Большинство древесных и кустарниковых псаммофитов, например песчаный саксаул (Haloxylon persicum) и солянка Рихтера (Salsola richteri), образуют мощные придаточные корни на стволах, погребенных песком. У некоторых деревянистых псаммофитов, например у песчаной акации (Ammodendron conollyi), на оголенных корнях образуются придаточные почки, а затем и новые побеги, которые позволяют продлить жизнь растения при выдувании песка из-под его корневой системы. У ряда травянистых псаммофитов образуются длинные корневища с острыми концами, которые быстро растут вверх и, достигнув поверхности, образуют новые побеги, избегая, таким образом, погребения.
Кроме того, у псаммофитов выработались в процессе их эволюции разнообразные приспособления у плодов и семян, направленные на обеспечение их летучести и способности перемещаться вместе с движущимся песком. Эти приспособления заключаются в образовании на плодах и семенах разнообразных выростов: щетинок - у джузгуна (Calligonum) и мешковидных вздутий - у осоки вздутой (Carex physodes), придающих упругость и легкость плодам; разнообразных летательных аппаратов.
К петрофитам (литофитам) относят растения, обитающие на каменистых субстратах - скальных обнажениях, каменистых и щебнистых осыпях, валунных и галечниковых отложениях по берегам горных рек. Все петрофиты являются так называемыми “пионерными” растениями, которые первыми заселяют и осваивают местообитания с каменистыми субстратами.
Топографические (орографические) факторы. Факторы рельефа оказывают на растения в основном косвенное действие, перераспределяя количество осадков и тепла по поверхности суши. В понижениях рельефа скапливаются выпавшие осадки, а также холодные массы воздуха, что является причиной поселения в этих условиях влаголюбивых и малотребовательных к теплу растений. Повышенные элементы рельефа, склоны южной экспозиции прогреваются лучше понижений и склонов другой ориентации, поэтому на них можно встретить более теплолюбивые и менее требовательные к влаге растения. Мелкие формы рельефа увеличивают разнообразие микроусловий, что создает мозаичность растительного покрова.
Особое влияние на распределение растений оказывает макрорельеф - горы, среднегорья и плоскогорья, которые создают на сравнительно небольшой площади значительные амплитуды высот. С изменением высоты меняются климатические показатели - температура и влажность, результатом чего является высотная поясность растительности. Горы часто являются барьером для проникновения растений из одних регионов в другие.
Биотические факторы. Большое значение в жизни растений имеют биотические факторы, под которыми подразумевают влияние животных, других растений, микроорганизмов. Это влияние может быть прямым, когда организмы, непосредственно соприкасаясь с растением, оказывают на него положительное или отрицательное воздействие (например, поедание травы животными), либо косвенным, когда организмы влияют на растение опосредованно, изменяя среду его обитания.
В жизни растений велика роль животного населения почвы. Животные размельчают и переваривают остатки растений, разрыхляют почву, обогащают почвенный слой органическими веществами, т. е. изменяют химизм и структуру почвы. Это создает условия для преимущественного развития одних растений и угнетения других. Насекомые и некоторые птицы опыляют растения. Известна роль зверей и птиц как распространителей семян и плодов растений.
Влияние животных на растения иногда проявляется через целую цепь живых организмов. Так, резкое снижение численности хищных птиц в степях ведет к быстрому размножению мышей-полевок, которые питаются зеленой массой степных растений. Это, в свою очередь, приводит к снижению урожайности степных фитоценозов и количественному перераспределению видов растений внутри сообщества.
Отрицательная роль животных проявляется в вытаптывании и поедании растений.
Очень разнообразно влияние одних растений на другие. Здесь можно выделить несколько типов взаимоотношений.
1. При мутуализме растения в результате совместного существования получают взаимную пользу. Примером таких взаимоотношений могут служить микориза, симбиоз клубеньковых бактерий-азотфиксаторов с корнями бобовых.
2. Комменсализм - это такая форма взаимоотношений, когда совместное существование для одного растения выгодно, а для другого безразлично. Так, одно растение может использовать другое в качестве субстрата (эпифиты).
3. Паразитизм - такая форма взаимоотношений, когда растение-паразит живет за счет растения-хозяина.
4. Конкуренция - проявляется у растений в борьбе за условия существования: влагу, питательные вещества, свет и т. п. Различают внутривидовую конкуренцию (между особями одного и того же вида) и межвидовую (между особями разных видов).
Антропические (антропогенные) факторы. Человек с давних времен оказывает влияние на растения, особенно заметно оно проявляется в наше время. Это влияние может быть прямым и косвенным.
Прямое воздействие - это вырубка леса, сенокошение, сбор плодов и цветов, вытаптывание и т. п. В большинстве случаев такая деятельность отрицательно сказывается на растениях и растительных сообществах. Численность некоторых видов резко сокращается, некоторые могут полностью исчезнуть. Происходит существенная перестройка растительных сообществ или даже смена одного сообщества другим.
Не меньшее значение имеет косвенное воздействие человека на растительный покров. Оно проявляется в изменении условий существования растений. Так появляются рудеральные, или мусорные, местообитания, промышленные отвалы. Отрицательное влияние на жизнь растений оказывает загрязнение атмосферы, почв, вод индустриальными отходами. Оно приводит к исчезновению на определенной территории некоторых видов растений и растительных сообществ в целом. Естественный растительный покров изменяется и в результате увеличения площадей под агрофитоценозы.
В процессе своей хозяйственной деятельности человек должен учитывать все взаимосвязи в экосистемах, нарушение которых нередко влечет за собой непоправимые последствия.
Классификация жизненных форм растений.Экологические факторы влияют на растение не изолированно друг от друга, а во всей их совокупности. Приспособленность растений ко всему комплексу условий местообитания отражает жизненная форма. Под жизненной формой понимают группу видов, сходных по внешнему облику (габитусу), который обусловливается сходством основных морфологических и биологических признаков, имеющих приспособительное значение.
Жизненная форма растений является результатом приспособления к определенной среде обитания и вырабатывается в процессе длительной эволюции. Поэтому признаки, характерные для жизненной формы, закрепляются в генотипе и проявляются у растений в каждом новом поколении. При выделении жизненных форм учитываются различные биологические и морфологические признаки растений: форма роста, ритмы развития, продолжительность жизни, характер корневых систем, приспособления к вегетативному размножению и т. д. Поэтому жизненные формы растений называют также биоморфами.
Существуют разные классификации жизненных форм растений, не совпадающие с классификацией систематиков, основанной на строении генеративных органов и отражающей “кровное родство” растений. Сходную жизненную форму принимают в сходных условиях растения совсем не родственные, принадлежащие к разным семействам и даже классам.
В основу биоморфологических классификаций можно, в зависимости от цели, положить разные признаки. Одна из наиболее распространенных и универсальных классификаций жизненных форм растений была предложена датским ботаником К. Раункиером. Она основана на учете приспособления растений к перенесению ими неблагоприятных условий - низких осенне-зимних температур в районах с холодным климатом и летней засухи в жарких и сухих областях. Известно, что от холода и засухи у растений страдают прежде всего почки возобновления, а степень защищенности почек в значительной мере зависит от положения по отношению к поверхности почвы. Этот признак и был использован К. Раункиером для классификации жизненных форм. Он выделил пять крупных категорий жизненных форм, назвав их биологическими типами. Схематически эти категории п
Дата добавления: 2015-08-08; просмотров: 3650;