Репродуктивные процессы в цветке
В цветке последовательно протекают важнейшие процессы, обеспечивающие вначале бесполое размножение и образование очередного поколения гаметофитов, а затем и половое размножение, завершающееся образованием нового поколения спорофитов в виде зародышей, расположенных в семенах. Цветковые являются разноспоровыми растениями. Они образуют два типа спор: микро- и мегаспоры.
Клетки бесполого размножения – микроспоры, образуются в ходе процесса, называемого микроспорогенезом в микроспорангиях, которыми являются гнезда пыльника. В результате ряда последовательных митотических делений клеток археспория, то есть образовательной ткани пыльника, возникают материнские клетки микроспор (микроспороциты), содержащие, как и все растение, диплоидный набор хромосом. Далее каждая материнская клетка в результате мейотического деления образует тетраду гаплоидных микроспор. Сформировавшаяся микроспора имеет оболочку и единственное ядро.
В ходе превращения микроспоры в пыльцевое зерно происходит два митотических деления и под защитой оболочки микроспоры возникают сначала две, далее три клетки – редуцированный мужской гаметофит. После формирования такого редуцированного гаметофита трехклеточная структура называется пыльцевым зерном (мужской особью). Пыльцевое зерно в начале развития содержит две клетки: маленькую генеративную и более крупную, часто называемую вегетативной. Микрогаметогенез крайне упрощен. Генеративная клетка делится однократно и из нее образуются две безжгутиковые мужские гаплоидные гаметы – спермии. В таком состоянии пыльцевое зерно готово к половому размножению. При попадании на рыльце пестика цветка вегетативная клетка пыльцевого зерна удлиняется и становится так называемой «пыльцевой трубкой», с мощью которой уже в семязачатке завершается оплодотворение (рис. 13.13).
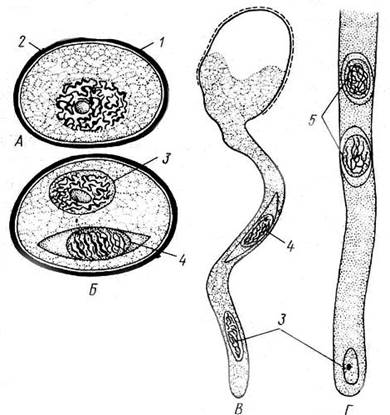
Рис. 13.13. Микроспора, пыльцевое зерно и его прорастание . А – микроспора; Б – пыльцевое зерно; В – формирование пыльцевой трубки; Г– часть пыльцевой трубки: 1 – экзина, 2 – интина, 3 – вегетативная клетка, дающая начало пыльцевой трубке, 4 – генеративная клетка, 5 – спермий.
Зрелое пыльцевое зерно окружено сложно устроенной оболочкой. Оболочка, часто называемая спородермой, состоит из двух главных слоев: внешнего, более толстого – экзины, и относительно тонкого внутреннего – интины. Экзина, содержащая особое высокомолекулярное вещество спорополленин, характеризуется необычайной стойкостью: она не растворяется в кислотах и щелочах, выдерживает температуру до 300 °С и сохраняется в фоссилизованном (окаменевшем) состоянии миллионы лет в геологических отложениях. Это позволяет вести спорово-пыльцевой анализ почв, донных отложений и осадочных толщ, определять, какие виды росли в разные геологические периоды. Экзина микроспор имеет на поверхности разнообразные скульптурные утолщения.
Особенности скульптуры важны при морфологической характеристике пыльцевых зерен отдельных таксонов. Наиболее развиты такие утолщения у энтомофильных растений. Интина менее стойка, чем экзина. Она состоит из целлюлозы и пектина и как бы облегает содержимое пыльцевого зерна.
Перенос пыльцевых зерен из тычинок на рыльца пестиков называется опылением. Для того чтобы образовался зародыш семени, должны произойти опыление и оплодотворение. Опыление впервые появляется у голосеменных, однако наибольшее разнообразие механизмов опыления и его совершенство достигаются у цветковых. Различают два типа опыления – самоопыление, или автогамию (от греческого «аутос» – сам), и перекрестное опыление, или ксеногамию (от греческого «ксенос» – чужой, «гамос» – брак). При самоопылении рыльце опыляется пыльцой того же цветка или пыльцой других цветков этой особи растения – гейтоногамия (девственное опыление). Обычно самоопыление осуществляется в распустившихся цветках, но иногда происходит в цветках закрытых, нераспустившихся (клейстогамных). В генетическом отношении все эти способы вполне равноценны.
Если перенос пыльцы осуществляется между цветками разных особей, то происходит перекрестное опыление. Оно свойственно не менее 90% видов цветковых растений. Перекрестное опыление обеспечивает обмен генами, поддерживает высокий уровень гетерозиготности популяций, определяет единство и целостность вида. Это создает широкое поле для деятельности естественного отбора. Строгое самоопыление встречается относительно редко (например, у гороха) и может вести к расщеплению вида на ряд чистых линий, то есть делает популяции гомозиготными. Иногда это приводит к затуханию микроэволюции. Но самоопыление способствует изоляции вновь возникших в результате мутаций форм, обособляя и фиксируя их в чистых линиях. По-видимому, для эволюционного процесса оптимально сочетание самоопыления и перекрестного опыления, что чаще всего и имеет место в природе. Однако преобладает перекрестное опыление и поэтому у цветковых обычно есть специальные приспособления морфологического и физиологического характера, предотвращающие или ограничивающие самоопыление. К ним относятся двудомность, дихогамия, гетеростилия, самонесовместимость. Наиболее надежное средство предотвращения самоопыления – двудомность, но при этом часть особей популяций (мужские особи) не дает семян. Однодомность устраняет автогамию, но не предохраняет от генетически равноценной ей гейтоногамии. Функциональная раздельнополость получила название дихогамии (от греческого «дихе» – на две части). Она проявляется в разновременном созревании пыльцы и рылец в цветках одного и того же растения. Дихогамия чаще всего встречается в форме протандрии. В этом случае раньше вызревает пыльца. При протогинии (другой форме дихогамии) происходит более раннее созревание рылец и завязей, в которых заключены семязачатки.
Иногда бывает полная физиологическая самонесовместимость. Она выражается в подавлении при самоопылении прорастания пыльцы на рыльце пестика той же особи. Самонесовместимость встречается у покрытосеменных более широко, чем двудомность. Она зарегистрирована более чем у 10000 видов цветковых.
Явление гетеростилии состоит в том, что у некоторых видов имеются две или даже три формы цветков (находящиеся на разных особях), различающиеся по длине столбиков и тычиночных нитей. В силу этого самоопыление у этих растений в значительной мере затруднено и дает обычно ничтожное количество семян. Классическими примерами растений, характеризующихся гетеростилией, служат виды первоцветов (Primula) и болотное растение дербенник иволистный (Lythrum salicaria).
Механизмы перекрестного опыления весьма разнообразны и являются главным объектом изучения особого раздела ботаники, называемого антэкологией. Их подразделяют на два основных типа – биотическое и абиотическое опыление. Биотическое опыление осуществляется животными, абиотическое – с помощью неживых факторов внешней среды. Наибольшее значение среди механизмов биотического опыления имеет энтомофилия (от греческого «энтомон» – насекомое, «филео» – любить). Насекомые сыграли выдающуюся роль в эволюции цветка. Цветки привлекают насекомых запасом пищи: пыльцой, нектаром. Для привлечения опылителей служат яркая окраска и характерный запах цветков; некоторое значение имеет их форма. Конкретные механизмы перекрестного опыления, осуществляемого насекомыми, очень разнообразны и связаны как со строением цветка, так и с особенностями тела насекомого. Видам некоторых семейств (бобовые, губоцветные, орхидные) свойственны особые способы опыления.
Помимо насекомых существенную роль в биотическом опылении, особенно в тропиках, играют птицы (орнитофилия), летучие мыши и некоторые нелетающие млекопитающие (например, крысы). Абиотическое опыление связано с переносом пыльцы ветром (анемофилия, от греческого «анемос» – ветер), реже (у болотных и водных растений) – водой. Анемофилы – это преимущественно растения открытых пространств. У анемофильных цветковых растений, как правило, невзрачный, сильно редуцированный околоцветник и крупные, часто мохнатые рыльца с огромной воспринимающей пыльцу поверхностью. Очень часто мелкие цветки анемофилов собраны в плотные или многоцветковые соцветия, пыльца их обильна, легка, а экзина почти лишена скульптурных утолщений, обычных для пыльцы энтомофильных растений.
Второй тип спор разноспоровых цветковых растений – мегаспоры образуются в процессе мегаспорогенеза, протекающего в семязачатке. Центральная часть семязачатка – нуцеллус, представляет собой мегаспорангий цветковых растений. Семязачаток состоит из диплоидных клеток, поскольку он является частью растения-спорофита. Одна из клеток нуцеллуса является материнской клеткой мегаспор. Она редукционно делится и формирует тетраду линейно расположенных гаплоидных клеток. (рис. 13. 14).
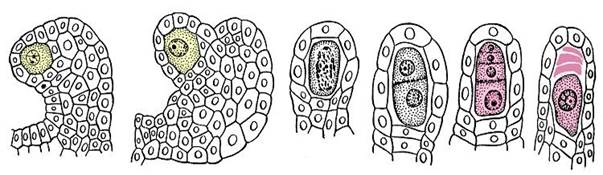
Рис. 13.14. Стадии развития мегаспор в семязачатке лобелии(Lobelia cardinalis)
Это и есть мегаспоры цветковых растений. Обычно три из четырех клеток-мегаспор в семязачатке отмирают, единственная жизнеспособная мегаспора развивается в женский гаметофит, или зародышевый мешок.
Формирование зародышевого мешка, получившее название мегагаметогенез, происходит путем трех последовательных делений ядра мегаспоры. В результате первого деления образуется двухъядерная клетка, которая сильно увеличивается в длину. Далее каждое из двух ядер делится еще дважды и возникает восьмиядерная клетка, имеющая по четыре ядра у каждого полюса. На этом деления завершаются. В центр клетки от каждого их полюсов перетекает по одному гаплоидному ядру и они сливаются в диплоидное вторичное ядро. В результате цитокинеза остальные ядра (по три у каждого полюса) формируют дочерние гаплоидные клетки. В итоге возникает семиклеточный зародышевый мешок. На микропилярном его конце обычно имеется гаплоидная яйцеклетка в окружении двух гаплоидных клеток-синергид. На халазальном конце расположены три гаплоидные клетки-антиподы. Основной объем зародышевого мешка составляет центральная диплоидная клетка (см. рис. 13.10).
Для осуществления оплодотворения необходимы два условия: зрелая жизнеспособная пыльца, попавшая на рыльце пестика, и сформировавшийся зародышевый мешок в семязачатке. Оплодотворению предшествует прорастание пыльцевого зерна. Оно начинается с разбухания зерна и выхода из апертуры пыльцевой трубки. В густой цитоплазме кончика пыльцевой трубки идут интенсивные физиологические процессы, вследствие которых несколько размягчаются ткани рыльца и столбика, в которые внедряется пыльцевая трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки и оба спермия. В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий (нуцеллус) через микропиле семязачатка, реже – иным образом. Проникнув в зародышевый мешок, пыльцевая трубка разрывается (под действием разницы осмотического давления в ней и давления в нуцеллусе) и ее содержимое изливается в зародышевый мешок. Один из спермиев сливается с яйцеклеткой и образуется диплоидная зигота, дающая затем начало зародышу. Второй спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка, что приводит к образованию триплоидного ядра, развивающегося затем в специальную питательную ткань – эндосперм (от греческого «эндон» – внутри, «сперма» – семя). Весь этот процесс получил название двойного оплодотворения. Он впервые описан в 1898 г. выдающимся русским цитологом и эмбриологом С.Г.Навашиным. Прочие клетки зародышевого мешка (антиподы и синергиды) разрушаются при проникновении пыльцевой трубки. Однако при гибели яйцеклетки они могут выполнить ее функцию.
Биологический смысл двойного оплодотворения, в отличие от голосеменных, у которых гаплоидный эндосперм развивается независимо от процесса оплодотворения, состоит в том, что у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа семян этим достигается существенная экономия энергетических и пластических ресурсов.
У многих цветковых (около 10% видов) в процессе эволюции половое размножение замещается различными формами бесполого. Из них наиболее известен апомиксис. Семена у апомиктичных растений образуются без оплодотворения. Апомиксис исключает генетическое расщепление, поэтому апомиктические формы образуют клоны, в пределах которых все особи имеют одинаковую генетическую и соматическую конституцию. Хорошим примером растения с апомиктическим образованием семян служит одуванчик с его поразительно высокой жизнеспособностью. Нередко у многолетних корневищных растений преобладает вегетативное размножение, а семенное ограничено.
Дата добавления: 2015-08-08; просмотров: 2431;