Двигательная иннервация
Формирование нервно-мышечного синапса.Мотонейроны направляют свои аксоны в места закладки скелетных мышц приблизительно в то время, когда появляются мышечные трубочки. Несколько аксонов подрастают к месту первого контакта (полинейронная иннервация), однако, только один наиболее активный аксон сформирует дефинитивный нервно-мышечный синапс (Рис. 7-10). В мышечной трубочке до контакта с аксоном мотонейрона экспрессируется эмбриональной формы н-холинорецепторов, состоящих из 2 –СЕ, 1 -, 1 - и 1 –СЕ. В месте вхождения аксонов в формирующуюся мышцу образуются первые неспециализированные нервно-мышечные контакты. Эмбриональные н-холинорецепторы, диффузно распределённые в мембране мышечной трубочки, концентрируются в области будущего синапса (рис. 7-11). Импульсация мотонейрона, передающаяся на мышечную трубочку, ингибирует экспрессию эмбриональной –СЕ холинорецептора. При этом из нервной терминали секретируются нейрогенные молекулы агрин и нейрегулины, в том числе ARIA [AChR (acetylcholinereceptor) inducing activity — индуцирующий активность холинорецепторов]. ARIA образуется в результате альтернативного сплайсинга первичного транскрипта гена nrg-1, транспортируется по аксону и секретируется в синаптическую щель. В мембране мышечной трубочки ARIA связывается с рецепторными тирозин киназами ErbB3 и ErbB4, принадлежащим к семейству рецепторов эпидермального фактора роста (EGF). Фосфорилированные рецепторы ErbB3 и ErbB4 инициируют фосфатидил инозитол 3-киназный (PI 3-K) и митоген-активированный протеин киназный (MAP-K) сигнальные каскады, активирующие экспрессию гена холинорецептора AChR в ядрах мышечной трубочки. Молекула агрина взаимодействует с мышечно-специфической тирозин киназой MuSK, обеспечивающей кластеризацию рапсина. Одна молекула рапсина связана с одной молекулой холинорецептора, при этом рапсин взаимодействует с белками цитоскелета постсинаптической мембраны, в том числе с утрофином, дистрогликанами, дистробревином, синтрофином, обеспечивая кластеризацию холинорецепторов в постсинаптической мембране (рис. 7-12).
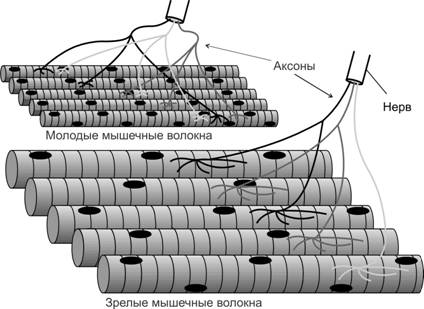
Рис. 7-10. Элиминация полинейронной иннервации. В плодном периоде мышечные волокна иннервируются 2-6 аксонами. В постнатальном периоде конкурентное взаимодействие нервных терминалей (в т.ч. за нейротрофические факторы, продуцируемые мышечными волокнами) устраняет неактивные аксоны и только один наиболее активный аксон сформирует дефинитивный нервно-мышечный синапс. [74]
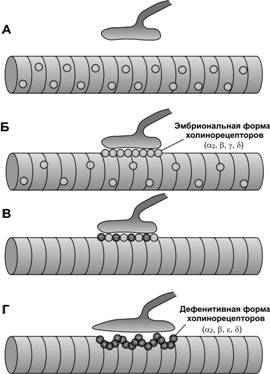
Рис. 7-11. Образование нервно-мышечного синапса. А. В мышечных трубочках ещё до установления контакта с аксоном экспрессируется эмбриональная форма холинорецепторов (2), которые равномерно распределены по плазмолемме миотубы. Б. Через несколько часов после подрастания аксона к мышечной трубочке происходит агрегация холинорецепторов в месте контакта аксона и миотубы.В. Через двое суток в результате электрической активности аксона ингибируется экспрессия СЕ холинорецепторов и начинается синтез СЕ. Г. В постнатальном периоде под нервной терминалью образуются складки плазмолеммы и агрегаты из ядер мышечного волокна. [74]
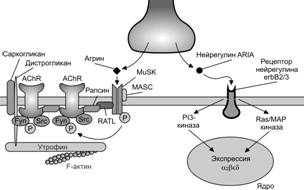
Рис. 7-12. Роль аксона в созревании постсинаптической мембраны. Из нервной терминали в синаптическую щель секретируются нейрогенные молекулы. Нейрегулин (ARIA) стимулирует экспрессию генов дефинитивной формы холинорецепторов (2), а агрин, активируя MuSK,способствует кластеризации рапсина, взаимодействующего с холинорецепторами и белками цитоскелета постсинаптической мембраны. [74]
Нейромоторная (двигательная) единица.Одна нейромоторная единица включает один мотонейрон и группу иннервируемых им экстрафузальных мышечных волокон. Количество и размеры двигательных единиц в различных мышцах значительно варьируют. Аксон одного из мотонейронов может иннервировать от одного до нескольких сотен мышечных волокон. Поскольку при сокращении фазные мышечные волокна подчиняются закону всё или ничего, то сила, развиваемая мышцей, зависит от количества активируемых (т.е. участвующих в сокращении мышечных волокон) двигательных единиц. Существует три типа нейромоторных единиц: медленная (S), быстрая неутомляемая (FR) и быстрая утомляемая (FF). Каждая нейромоторная единица образована только медленносокращающимися, быстросокращающимися неутомляемыми или быстросокращающимися утомляемыми мышечными волокнами. Тип нейромоторной единицы зависит от частоты импульсации, генерируемой мотонейроном. Различают медленные (S) и быстрые мотонейроны (F) с частотой импульсации 1525 Гц и 4060 Гц, соответственно. S-мотонейроны иннервируют медленносокращающиеся мышечные волокна, а F-мотонейроны — быстросокращающиеся. Мышечные волокна, принадлежащие разным нейромоторным единицам, имеют мозаичное распределение в скелетной мышце (рис. 7-13).
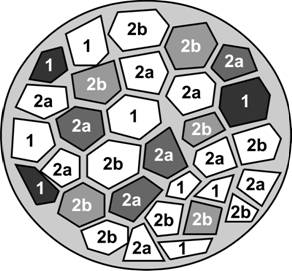
Рис. 7-13. Мозаичное распределение мышечных волокон разных нейромоторных единиц. Мышечные волокна одного типа (1, 2a, 2b), принадлежащие одной нейромоторной единице, находятся между волокон других нейромоторных единиц. [77]
Полинейронная иннервация. В постнатальном периоде фазное мышечное волокно иннервируются только одним мотонейроном, в то же время тонические мышечные волокна, например наружных глазных мышц, получают полинейронную иннервацию.
Дата добавления: 2015-07-07; просмотров: 2259;