Сигнальные молекулы.
Основные задачи регуляции метаболизма и клеточных функций:
1 внутриклеточное и межклеточное согласование клеточных процессов.
2. исключение «холостых» циклов метаболизма, продукты которых не востребованы,
3. эффективное образование и использование энергии,
4. поддержание гомеостаза,
5. приспособление организма к условиям окружающей среды.
Регуляция метаболизма: внутренняя и внешняя. Внутренняя регуляция - управляющие сигналы образуются и действуют внутри одной и той же клетки (само-регуляция). Внешняя регуляция — управляющие сигналы поступают к клетке из внешней среды
Внутренняя регуляция осуществляется путем изменения активности ферментов активаторами или ингибиторами. Особенно активно при внутренней регуляции работают аллостерические ферменты (ФОСФОРИЛАЗА}
Внешняя регуляция обеспечивается специализированными сигнальными молекулами - эндогенные химические соединения, которые в результате взаимодействия с ферментами, обеспечивают внешнее управление биохимическими процессами в клетках-мишенях.
Клетка-мишень — это клетка, имеющая специализированные воспринимающие рецепторы для данного вида сигнальных молекул. Сигнальные молекулы являются лигандами для рецепторов клеток-мишеней.
Характерные особенности сигнальных молекул:
1. малый период жизни (динамичность, оперативность регуляции),
2. высокая биологическая активность (действие развивается при очень низких концентрациях).
3. уникальность, неповторимость действия. Эффекты одного типа сигнальных молекул не могут быть смоделированы другим. Это обеспечивает разнообразие регуляции.
4. наличие эффекта усиления (одна сигнальная молекула может усиливать каскады биохимических реакций).
5. один вид сигнальных молекул может иметь несколько клеток-мишеней.
6. реакция разных клеток-мишеней на одну и ту же сигнальную молекулу отличается (объясняется многообразием рецепторов и их своеобразием).
Способы внешнего управления клетками-мишенями:
· Управление экспрессией генов (биосинтез белков и ферментов). Это медленный способ регуляции.
· Управление активностью ранее синтезированных белков:
· управление активностью ферментов, следовательно, изменение биохимических процессов.
· изменение активности функциональных белков, следовательно, прямое изменение функции клеток. Например, влияние сигнальных молекул на ионный канал вызывает деполяризацию мембраны и формирование потенциала действия. Чрезвычайно быстрый эффект.
Виды регуляторных эффектов сигнальных молекул:
1 Эндокринный. Сигнальные молекулы поступают с током крови из желудочно-воротной системы к клеткам-мишеням. Так действует большинство гормонов.
2. Паракринный - сигнальные молекулы вырабатывают в пределах одного органа или участка ткани. Таким образом действуют большинство факторов роста.
3. Аутокринные - сигнальные молекулы действуют на клетку, их образовавшую.
КЛАССИФИКАЦИЯ СИГНАЛЬНЫХ МОЛЕКУЛ
По химической природе:
1. Органические (производные аминокислот, жиров) СТЕРОИДЫ, ПРОСТОГЛАНДИНЫ.
2. Неорганические 1992г. МОНООКСИДАЗОТА (N0).
По физико-химическим свойствам:
1. Липофобные не могут проникать через мембрану клетки. Они растворимы в воде.
2. Липофильные растворяются в жирах. Свободно проникают через ЦПМ и действуют на рецепторы внутри клетки. Например, производные холестерина: МИНЕРАЛО -, ГЛЮКОКОРТИКОИДЫ, ЭСТРОГЕНЫ, АНДРОГЁНЫ, ЙОДТИРОНИНЫ, N0.
Но биологическому принципу:
1. Гормоны - сигнальные молекулы с выраженным эндокринным эффектом.
2. Цитокины - факторы роста. Это сигнальные молекулы белковой природы, которые выделяются неспециализированными клетками организма. Они регулируют рост, дифференцировку, пролиферацию соседних клеток. Действие пара - и аутокринно.
3. Нейромедиаторы — сигнальные молекулы, вырабатывающиеся нервными клетками, координирующие работу нейронов и управление периферическими тканями. Их действие связано с влиянием на ионные каналы. Они изменяют их проницаемость и вымывают деполяризацию мембраны.
МЕХАНИЗМ ДЕЙСТВИЯ СИГНАЛЬНЫХ МОЛЕКУЛ.
Механизм действия ОРГАНИЧЕСКИХ ЛИПОФИЛЬНЫХ сигнальных молекул.
1. взаимодействие с внутриклеточными рецепторами,
2. регуляторный эффект связан с изменением количества белков в результате влияния на экспрессию генов (действуют через геном).
3.. биологическое действие продолжительное, но развивается медленно в пределах часов.
Факторы, необходимые для их действия:
1. сигнальные молекулы.
2 воспринимающий внутриклеточный рецептор, связанный с шапероном,
3. участок ДНК, регулирующий транскрипцию определённых генов (ЭНХАНСЕР, САЙЛЕНСЕР),
4, белок синтетический аппарат клетки.
Этапы действия:
1. проникновение внутрь клетки,
2. связывание с внутриклеточным рецептором,
3. освобождение шаперона (запуск таймера действия),
4. взаимодействие комплекса сигнальных молекул с регуляторными элементами ДНК, изменение биосинтеза некоторых белков, в том числе и их ферментов
5. изменение метаболизма и клеточных функций.
Механизм прекращения действия органических липофильных сигнальных молекул:
1. разрушение рецепторов, обусловленное отсутствием защиты со стороны шаперона,
2. протеолиз синтезированных белков,
3.разрушение факторов транскрипции, участвующих в передаче сигналов к структурному гену.
По перечисленному механизму действуют СТЕРОИДНЫЕ гормоны и ИОДТИРОНИН.
Механизм действия ЛИПОФОБНЫХ сигнальных молекул:
1. взаимодействие с поверхностными рецепторами,
2. сигнал передаётся от рецептора внутрь клетки (ТРАНСДУКЦИЯ) и устанавливается с помощью внутриклеточных регуляторов: высоко- и низкомолекулярных.
Высокомолекулярные регуляторы - это регуляторные белки. Они опосредуют действие сигнальной молекулы внутри клетки.
Низкомолекулярные регуляторы небелковой природы. Его называют второй МЕССЕНДЖЕР (первый МЕССЕНДЖЕР - сама сигнальная молекула) - полноправный представитель сигнальной молекулы внутри клетки. Это ионы кальция, ДИАЦИЛГЛИЦЕРОЛ, ИНОЗИТОЛТРИФОСФАТ, цАМФ и цГМФ.
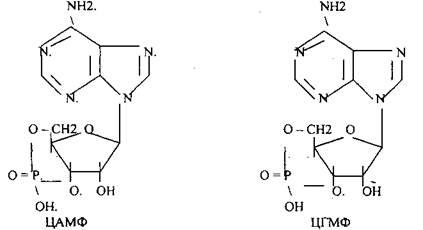
3. биологическое действие обусловлено сочетанием регуляции активности ранее синтезированных белков и регуляция экспрессии генов. Регуляторный эффект двухфазный: первая фаза быстрая, но не продолжительная, она обеспечивает изменение структуры и активности ферментов; вторая фаза медленная за счёт изменения количества ферментов.
МЕХАНИЗМ ДЕЙСТВИЯ, ЗАВИСИМЫЙ ОТ ЦАМФ.
Факторы, необходимые для этого:
· растворимая в воде сигнальная молекула;
· поверхностные рецепторы клетки-мишени;
· внутриклеточный трансдуктор G-белок. Состоит из 3 единиц: альфа, бета, гамма.
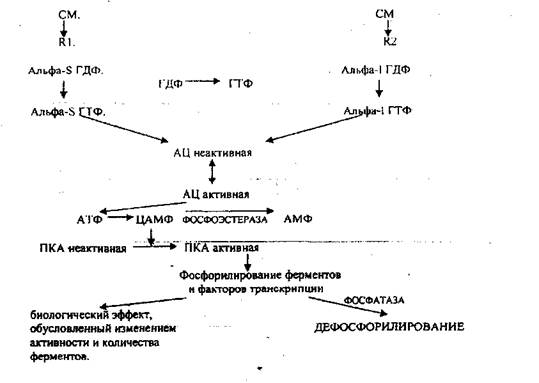
G-белок может быть ингибирующий и активирующий. Он находится в непосредственной близости к рецептору. При взаимодействии сигнальной молекулы с рецептором он активируется, его активность обусловлена альфа единицей. Она оказывает влияние на внутриклеточный фермент - АДЕНИЛАТЦИКЛАЗУ (превращает АТФ в ЦАМФ). Альфа-5 повышает активность, альфа-1 понижает активность G-белка. G-белок способен присоединять ГДФ или ГТФ. Альфа-единица активна, когда связана с ГТФ.
· АДЕНИЛАТЦИКЛАЗА (АЦ);
· ПРОТЕИНКИНАЗА-А ЦАМФ-зависимая. Она катализирует реакцию фосфорилирования белков. В результате белки изменяют активность;
· Регуляторные элементы ДНК (ЭНХАНСЕР и САЙЛЕНСЕР);
· ФОСФОДИЭСТЕРАЗА - разрушает ЦАМФ;
· ФОСФАТАЗА - дефосфорилирует белки;
· Белок - синтетический аппарат клетки.
Этапы, стимулирующие ЦАМФ - зависимый механизм:
1. взаимодействие сигнальной молекулы с рецептором;
2. изменение конформации G-белка;
3. замена ГДФ на ГТФ в альфа - S единице G-белка;
4. альфа-S ГТФ активирует АЦ;
5. АЦ синтезирует ЦАМФ;
6. ЦАМФ активирует ПРОТЕИНКИНАЗУ-А (ПКА);
7. ПКА фосфорилирует белки и белковые факторы транскрипции, изменяющие активность и количество ферментов;
8. Прекращение действия, если рецептор освободился - альфа-S-G-ГТФ-азная активность:
альфа-единица может разрушать ГТФ - это таймер действия.
9. ФОСФОДИЭСТЕРАЗА - разрушает ЦАМФ.
10. ФОСФАТАЗА - ДЕФОСФОРИЛИРУЕТ белки.
Этапы, ингибирующие ЦАМФ - зависимый механизм:
С первого по третий те же самые этапы, отличие в G-белке (альфа-1 единица). Четвёртый этап - связывание ГТФ с альфа-1 единицей будет ингибировать АЦ. Ингибирующий механизм противодействует и прекращает эффекты ЦАМФ в клетке.
ЦГМФ - зависимый стимулирующий механизм действия.
Рецептор встроен в мембрану клетки и связан с ферментом ГУАНИЛАТЦИКЛАЗОЙ (ГЦ). При присоединении сигнальной молекулы ГЦ активируется и катализирует реакцию ГТФ----> ЦГМФ. Последний активирует ПРОТЕИНКИНАЗУ G (ПКG), а она запускает реакцию фосфорилирования белков (ферментов и факторов транскрипции).
ТИРОЗИНКИНАЗНЫЙ МЕХАНИЗМ ДЕЙСТВИЯ.
ТИРОЗИНКИНАЗА - фермент, фосфорилирующий белки. По этому механизму действует большинство факторов роста и пролиферации. Рецептор представлен дуплетом, который после взаимодействия с сигнальной молекулой ДИМЕРАЛИЗУЕТСЯ, что запускает АУТОФОСФОРИЛИРОВАНИЕ остатков тирозина в центральной части рецептора. Наблюдается отсутствие МЕССЕНДЖЕРОВ. Рецептор оказывает влияние на ферментные системы клетки. Ом может поступать в ядро вместе с сигнальными молекулами и усиливать транскрипцию генов и изменять митотическую активность клетки.
Механизм действия НЕОРГАНИЧЕСКИХ ЛИПОФОБНЫХ сигнальных молекул (NO).
NO беспрепятственно проникает через мембрану клетки. Это короткоживущая молекула, образующаяся в организме под действием NO-СИНТАЗЫ из аминокислоты АРГ. В клетке NO взаимодействует с ГЦ, активирует её, что вызывает накопление в клетке ЦГМФ, который активирует ПРОТЕИНКИНАЗУ-G, и развивается клеточный ответ по выше рассмотренному механизму. Эффекты NO:
· фактор расширения сосудов;
· регулятор АПОПТОЗА (запрограммированной клеточной смерти);
· NO является свободным радикалом, поэтому он способен влиять на процессы перекисного
окисления жиров и регулирования функций МИТОХОНДРИЙ;
· является ИММУНОМОДУЛЯТОРОМ.
1. РОЛЬ ГИПОТАЛАМУСА В РЕГУЛЯЦИИ ОБМЕНА ВЕЩЕСТВ.
2. ГОРМОНЫ ГИПОФИЗА.
ГИПОТАЛАМУС является компонентом и своеобразным «выходным каналом» лимбической системы. Это отдел промежуточного мозга, контролирующий различные параметры гомеостаза. С одной стороны он связан с ЦНС (центры ВНС), с другой с гипофизом через нервные проводники и особую портальную систему.
ГИПОТАЛАМУС участвует во многих функциях нервной регуляции, выделяя НЕЙРОТРАНСМИТТЕРЫ и НЕЙРОМОДУЛЯТОРЫ, а также регулирует эндокринную систему.

Многие из факторов ГИПОТАЛАМУСА образуются в периферических тканях (ТРГ, КРГ, СС). СС больше всего в D-клетках ПЖЖ. Действуют через ЦАМФ (АДЕНИЛАТЦИКЛАЗУ). Гормоны ГИПОТАЛАМУСА стимулируют транскрипцию генов гормонов гипофиза.
ГИПОФИЗ.
В нём выделяют три доли;
1 Передняя } АДЕНОГИПОФИЗ
2 Промежуточная (выделяет тропные гормоны)
3. Задняя - НЕЙРОГИПОФИЗ.
ТРОПНЫЕ гормоны можно разделить на 3 группы в зависимости от их химической природы:
· простые белки,
· ГЛИКОПРОТЕИНЫ;
· ПЕПТИДЫ, образующиеся из предшественников ПРООПИОМЕЛАНОКОРПИНА (ПОМК).
ПРОСТЫЕ БЕЛКИ.
К ним относятся: ГР, ПРЛ, ПЛАЦЕНТАРНЫЙ ХОРИОНИЧЕСКИЙ СОМАТОМАММОТРОПИН (ХС). Состоят из 191 - 198 аммиокислот ГР близок по строению с ХС на 85%, ПРЛ - на 35%.Среди этих гормонов наибольшую концентрацию имеют ГР. Секреция ГР эпизодическая. Максимальная секреция в момент засыпания или перед пробуждением. На секрецию влияют стресс, голод, физические упражнения. ГР сберегает глюкозу для тканей.
Он повышает ЛИПОЛИЗ— > повышается концентрация жирных кислот—> повышает транспорт в клетку-----> повышает концентрацию аминокислот -----> повышает ГЛЮКОНЕОГЕНЕЗ— > повышение глюкозы — — > мозг
ГР действует через инсулиноподобный фактор рост (ИФР), близкий по строению к проинсулину. Выделяют ИФР 1 и ИФР 2, но главным посредником действия ГР является ИФР 1.
Влияние ГР на обмен веществ:
1. Стимуляция синтеза белка:
повышение транспорта аминокислот в клетки, через ИФР 1.
повышение интенсивности процесса трансляции, через ИФР 1.
повышение синтеза РНК и ДНК
2. Влияние на углеводный обмен:
- повышение глюкозы (уменьшение утилизации глюкозы периферическими тканями, стимуляция процессов ГЛЮКОНЕОГЕНЕЗА),
3.Влияние на минеральный обмен (через ИФР1) задержка кальция, фосфора, магния в организме,
4. Влияние на липидный обмен (не через ИФР 1):
повышение ЛИПОЛИЗА,
повышение окисления ВЖК.
ГИПОСЕКРЕЦИЯ в детском возрасте приводит к нарушению синтеза белка, понижению минерализации костной ткани, задержке роста и развитию карликовости в нескольких вариантах,
ГИПЕРСЕКРЕЦИЯ гормона роста в детском возрасте приводит к развитою гигантизма, во взрослом развитии акромегалии. Нарушение синтеза гормона роста на умственную деятельность не влияет.
ПРЛ состоит из 198 аминокислот. Синтезируется при беременности и лактации. Функция заключается в инициации и поддержании лактации.
ХС проявляет ЛАКТОГЕННУЮ и МОТЕОТРОПНУЮ активность. Функция заключается в регуляции синтеза молока, развития желтых тел и развития плода.
ГЛИКОПРОТЕИНЫ
К ним относятся ТТГ, гонадотропины (ЛГ, ФСГ, ПЛАЦЕНТАРНЫЙ ХОРИОНИЧЕСКИЙ ГОНАДОТЮПИН (ХГЧ)). Действуют через ЦАМФ.
ФСГ:
Женские клетки-мишени - фолликулярные клетки яичника,
Мужские клетки-мишени - клетки СЕРТОЛИ семенников,
В женском организме стимулирует рост фолликулов, подготавливают их к действию ЛГ. В мужском организме индуцирует синтез АНДРОГЕНСВЯЗЫВАЮЩЕГО белка. Стимулирует рост семенных канальцев семенников и сперматогенез.
ЛГ:
Мужские клетки-мишени - клетки желтых тел.
Женские клетки-мишени - клетки ЛЕЙДЕГА.
Стимулирует образование в женском организме ПРОГЕСТЕРОНА, а в мужском ТЕСТОСТЕРОНА. Предшественником их является ХОЛЕСТЕРОЛ. ЛГ индуцирует овуляцию у женщин, ХГЧ синтезируется в плаценте и близок по эффектам к ЛГ. Появляется в моче после имплантации зародыша. Его определение служит диагностическим маркером беременности.
ТТГ:
Клетки-мишени - ТИРЕОЦИТЫ.
Повышает синтез ТРИЙОДТИРОНИНА и ТИРОКСИНА.
Повышает гидролиз белка ТИРЕОГЛОБУЛИНА.
Повышает включение йода в структуру ТИРЕОИДНЫХ гормонов.
Стимулирует синтез белка и нуклеиновых кислот в щитовидной железе (способствует повышению количества и росту ТИРЕОИДНЫХ гормонов)
ПЕПТИДЫ СЕМЕЙСТВА ПОМК.
ПОМК белок, состоящий из 285 аминокислот. Процессинг его происходит в передней и промежуточной доле гипофиза. Из него образуются пептиды: АКТГ, бета ЛИПОТРОПИН, N-концевой пептид.
АКТГ:
1. альфа - МЕЛАНОЦИТСТИМУЛИРУЮЩИЙ гормон (МСГ) (промежуточная доля).
2. КОРТИКОТРОПИНОПОДОБНЫЙ ПЕПТИД
Бета ЛИПОТРОПИН:
1. бета ЭНДОРФИНЫ: альфа - ЭНДОРФИНЫ, гамма - ЭНДОРФИНЫ.
2. Альфа - ЛИПОТРОПИН,
3 бета - МСГ.
N-концевой ПЕПТИД: гамма - МСГ
АКТГ – полипептид, состоящий из 39 аминокислот. Клетки-мишени - клетки коры надпочечников. Стимулирует синтез стероидов коры надпочечников из холестерина через ЦАМФ. Повышается АДЕНИЛАТЦИКЛАЗА в жировой ткани.
Усиливается ЛИПОЛИЗ ®ВЖК® АЦЕТИЛ-КОА® ХОЛЕСТЕРОЛ ® СТЕРОИДНЫЕ ГОРМОНЫ.
АКТГ стимулирует процессы пентозного цикла и является поставщиком НАДН2. Повышает синтез белка и нуклеиновых кислот в надпочечниках. ГИПЕРСЕКРЕЦИЯ проявляется в виде синдрома КУШИНГА:
· повышение пигментации,
· отрицательный азотистый баланс, выход азота, калия, фосфора.
· задержка в организме натрия (отёки, повышение АД).
Бета - ЛИПОТРОПИН усиливает липолиз.
Бета - ЭНДОРФИНЫ в гипофизе связаны и неактивны. В гипоталамусе и ЦНС они переходят в активную форму и играют роль нейромедиаторов, трансмиттеров. Понижают чувствительность к боли. Играют важную роль в эмоциональных состояниях.
МСГ стимулирует образование меланина.
ГОРМОНЫ ЗАДНЕЙ ДОЛИ ГИПОФИЗА
Вазопрессин (Антидиуретический гормон - АДГ), Окситоцин.
Образуются в ядрах гипоталамуса. Являются циклическими НАНОПЕПТИДАМИ (9 аминокислот)
АДГ: клетки-мишени клетки кровяных сосудов, дистальных извитых канальцев и собирательных трубочек почек. Повышают реабсорбцию воды в почках. Действует через ЦАМФ.
ГИПОСЕКРЕЦИЯ - несахарный диабет (10-12 л. мочи в сутки).
ОКСИТОЦИН: клетки-мишени - главные миоциты матки, клетки молочных желез. Повышает сокращение матки и секрецию молока. Используется для стимуляции родовой деятельности.
1. ГОРМОНЫ ЩИТОВИДНОЙ И ПАРДЩИТОВИДНОЙ ЖЕЛЕЗ.
2. ГОРМОНЫ ПЖЖ.
3 ГОРМОНЫ НАДПОЧЕЧНИКОВ,
ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ.
В щитовидной железе синтезируется и выделяется 3 гормона:
· ТРИЙОДТИРОНИН (ТЗ).
· ТЕТРАЙОДТИРОНИН (Т4).
· КАЛЬЦИТОНИН.
трийодтиронин и тетрайодтиронин синтезируются из аминокислоты тирозина, включённой в состав тиреоглобулина. Он подвергается йодированию с образованием 3-МОНОЙОДТИРОЗИНА, 3,5-ДИЙОДТИРОЗИНА, ТЕТРАЙОДТИРОНИНА.
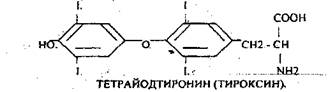
Через СООН- и NН2-группы тироксин встроен в структуру тиреоглобулина. Две молекулы тироксина связываются с помощью конденсирующего фермента. Для синтеза тироксина требуется йодид. Превращение йода в йодид происходит путём окисления с помощью ТИРЕОПЕРОКСИДАЗЫ. Синтез тиреоидных гормонов происходит в составе тиреоглобулина.
В свободном виде гормоны из железы выделяются в кровь после гидролиза тиреоглобулина с образованием свободных Т3 и Т4. Этот процесс стимулирует ТТГ гипофиза. Тормозит распад тиреоглобулина ионы йода, т.е. с одной стороны ионы йода необходимы для синтеза ТИРЕОГЛОБУЛИНА, с другой стороны при их избытке они тормозят его гидролиз, поэтому для лечения гипертириоза используют КI.
Т3 и Т4 плохо растворимы в воде и транспортируются с кровью тироксин связывающим глобулином (ТСГ). В клинической практике большой интерес представляет определение свободных ТЗ и Т4.
МЕХАНИЗМ ДЕЙСТВИЯ ТИРЕОИДНЫХ ГОРМОНОВ.
Т.к. они гидрофобны, то легко проникают через мембрану клетки. Рецепторы к ним находятся внутри клетки, главным образам в ядре, небольшая часть в цитоплазме. Сродство рецепторов к ТЗ больше, чем к Т4в 10 раз, несмотря на то, что образуется больше Т4. Рецептор внутри ядер негистоновый белок хроматина. ВЛИЯНИЕ НА МЕТАБОЛИЗМ
· Повышение поглощения тканями кислорода за исключением мозга, ретикуло-эпителиальной системы и гонад.
· Повышается активность натрии-калиевой АТФ-азы, при этом уровень атф понижается.
· Повышается синтез белка, положительный азотистый баланс, эффект физиологической концентрации гормонов. При повышении концентрации тиреоидных гормонов синтез белка понижается и наблюдается отрицательный азотистый баланс.
· Повышает транскрипцию гена гормона роста.
НАРУШЕНИЕ ФУНКЦИЙ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
ГИПОФУНКЦИЯ (ГИПОТИРЕОЗ):
1. У детей возникает кретинизм, проявляющийся:
В отставании в росте.
В отставании в физическом развитии;
В отставании в умственном развитии.
2. У взрослых возникает МИКСЕДЕМА (слизистый отёк):
понижается распад гликопротеинов;
повышение онкотического давления;
отёк тканей.
3.ЭНДЕМИЧЕСКИЙ ЗОБ возникает в регионах с недостатком йода в воде и пище. Понижается объём железистой ткани, но увеличивается объём самой железы за счёт соединительной ткани.
ГИПЕРФУНКЦИЯ (ГИПЕРТИРЕОЗ) проявляется:
1. Повышением потоотделения.
2. Экзофтальм.
3. Повышение окислительных процессов.
4. Повышение температуры тела.
5. Похудание и т.д.
КАЛЬЦИТОНИН - полипептид, синтезирующийся К-клетками щитовидной железы. Снижает уровень кальция в крови.
ГОРМОНЫ ПАРАЩИТОВИДНОЙ ЖЕЛЕЗЫ.
Синтезирует ПАРАТИРЕОИДНЫЙ ГОРМОН (паратгормон) ПТГ. Это пептид, состоящий из 84 аминокислотных остатков.
Увеличивает уровень кальция в крови за счёт:
1. стимулирует выход кальция и фосфора из костей в кровь.
2. повышает реабсорбцию кальция в почках,
3. стимулирует образование из витамина D3 гормона кальцитриола в почках, который
стимулирует всасывание кальция в кишечнике.
ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ.
ПЖЖ - это смешанная железа, осуществляющая эндокринную и экзокринную функцию. Эндокринная функция осуществляется островками ЛАНГЕРГАНСА. В них образуется 4 гормона четырьмя видами клеток:
· ИНСУЛИН (70%) синтезируется в В-клетках.
· ГЛЮКАГОГОН(25%) синтезируется в А-клетках,
· СОМАТОСТАТИН (5%) синтезируется в D-клетках.
· ПАНКРЕАТИЧЕСКИЙ ПОЛИПЕПТИД синтезируется в F-клетках.
Инсулин - это пептид, состоящий из 51 аминокислотного остатков в двух цепях.

ПРЕИСУЛИН состоит из 84 АК.
Процессинг: ПРОИНСУЛИН® ИНСУЛИН + С-ПЕПТИД
В клинике определяется количество инсулина и С-пептида.
МЕХАНИЗМ ДЕЙСТВИЯ ИНСУЛИНА.
Клетки-мишени: клетки печени, миоциты, жировой ткани.
Рецепторы находятся на поверхности ЦПМ, т.к. инсулин не может проникать в клетки без связи с рецептором. Может встречаться явление интернализации рецепторов, т.е. проникновение комплекса инсулин—рецептор в клетку путём эндоцитоза и последующее его разрушение. Т.о. количество рецепторов к инсулину снижается. С этим явлением связано возникновение инсулин независимого сахарного диабета.
ВЛИЯНИЕ НА МЕТАБОЛИЗМ:
1. Углеводный обмен: это единственный гормон, понижающий уровень глюкозы в крови, за счёт повышения транспорта глюкозы через мембрану клеток жировой и мышечной ткани. В печени глюкоза легко проникает в гепатоциты в силу разницы её концентрации в крови и в клетках. Но в печени инсулин повышает активность фермента ГЛЮКОКИНАЗЫ, катализирующего превращение глюкозы в глюкозо-6-фосфат. Инсулин повышает активность процессов гликолиза, повышает активность фермента ГЛЮКОГЕНСИНТЕТАЗЫ (синтез гликогена). Понижает активность глюкозо-6-фосфатазы, катализирующей превращение глюкозо-6-фосфата в глюкозу. Инсулин необходим для усвоения глюкозы
2. ЛИПИДНЫЙ ОБМЕН: ингибирует липолиз. Стимулирует липогенез из углеводов.
3. Обмен белков:
· Стимулирует синтез белков.
· Стимулирует транспорт аминокислот в клетку.
· Тормозит распад белков.
· Активизирует процесс транскрипции и трансляции.
· Стимулирует пролиферацию клеток за счет повышения секреции фактора роста фибробластов,
тромбоцитарного фактора роста, фактора роста эпидермиса. Дефицит инсулина проявляется в виде сахарного диабета.
1 Инсулин зависимый сахарный диабет связан с нарушением секреции инсулина (генетические нарушения, поражение ПЖЖ). Встречается у 10% больных.
2 Инсулин независимый сахарный диабет - уменьшение количества рецепторов к инсулину в результате интернализации. Встречается у 90% больных. Предрасполагающий фактор -ожирение.
· Клинические проявления:
· голод клеток на фоне ГИПЕРГЛИКЕМИИ,
· ПОЛИФАГИЯ,
· ПОЛИДИПСИЯ,
· ГЛЮКОЗУРИЯ (более 10ммоль/л),
· ПОЛИУРИЯ,
· КЕТОЗ - повышение кетоновых тел (за счёт не окисления АЦЕТИЛ-КОА),
· КЕТОАЦИДОЗ (диабетическая кома).
ГЛЮКАГОН полипептид, состоящий из 29 аминокислот.
МЕХАНИЗМ ДЕЙСТВИЯ.
Клетки - мишени: гепатоциты.
Рецепторы лежат на поверхности мембраны клеток. Действует через циклическую АМФ. Активизирует превращение фосфорилазы В в фосфорилазу А, в результате гликоген расщепляется с образованием глюкозы.
ВЛИЯНИЕ НА МЕТАБОЛИЗМ:
· Повышает концентрацию глюкозы в крови,
· Усиливает процессы глюконеогенеза.
· Повышает интенсивность липолиза.
СОМАТОСТАТИН - пептид, состоящий из 14 аминокислот. Подавляет образование других гормонов ПЖЖ.
Функция ПАНКРЕАТИЧЕСКОГО ПОЛИПЕПТИДА недостаточно изучена.
ГОРМОНЫ НАДПОЧЕЧНИКОВ.
1 . Мозговой слой
А) Адреналин (гормон) образуются из аминокислоты тирозина
В) Норадреналин (нейромедиатор).
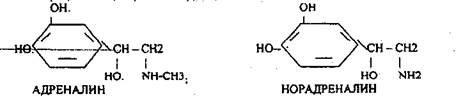
Это гормоны стресса. Их действие близко.
МЕХАНИЗМ ДЕЙСТВИЯ,
Клетки мишени: клетки печени, скелетных мышц, сердца, слюнных желез, матки.
Рецепторы находятся на поверхности мембран. Посредником является Ц АМФ, которая активизирует протеинкиназу.
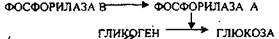
ВЛИЯНИЕ НА МЕТАБОЛИЗМ.
1. Повышает уровень глюкозы в крови.
2. Адреналин действует не только на печень, но и на мышцы, где из глюкозы образуется молочная кислота.
3. Усиливают липолиз.
4. Повышается содержание неэтерефицированных жирных кислот.
5. Повышает АД, частоту сердечных сокращений и т.д.
2.Корковый слой (более 30 стероидов)
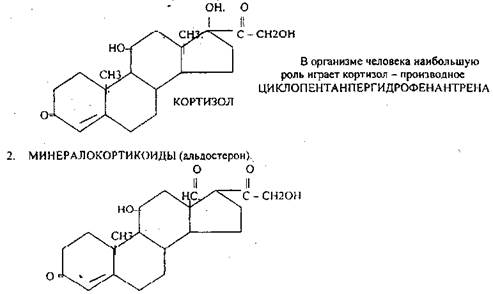
1.ГЛЮКОКОРТИКОИДЫ (кортикостерон, кортизол, кортизон).
МЕХАНИЗМ ДЕЙСТВИЯ.
Механизм действия цитозольный (проникают через мембрану). Рецепторы находятся внутри клетки. Действуют по принципу гормон—ген—белок. В печени, почках усиливают синтез белков; в лимфатической ткани, соединительной ткани, скелетных мышцах тормозят синтез белка.
ВЛИЯНИЕ НА МЕТАБОЛИЗМ.
1. Глюкоркортикоиды:
повышают глюконеогенез, повышают синтез гликогена, усиливают липолиз, усиливают образование кетоновых тел,
понижают синтез антител, следовательно, противовоспалительное и противоаллергическое действие.
2. Минералокортикоиды;
повышают транспорт натрия через мембрану, задерживают натрий и хлор в организме, понижают содержание калия в организме. Недостаточность гормонов коры ведёт к АДДИСОНОВОЙ болезни: пигментация кожи, - ГИПЕРКАЛИЕМИЯ, ГИПОТОНИЯ, понижение резистентности к стрессовым воздействиям.
Дата добавления: 2015-08-04; просмотров: 14254;