Физиологические теории мотиваций.
Первая группа теорий мотивации основана на представлении о сигнализации от периферических органов тела. В. Кеннон {1932, 1934), Д. Хебб (1949) полагали, что мотивации определяются стремлением организма избежать неприятных эмоциональных ощущений, сопровождающих различные побуждения. Животное утоляет жажду, чтобы избавиться от неприятной сухости в полости рта и глотки, поедает пищу, чтобы избавиться от мышечных сокращений пустого желудка, и т. д.
Дальнейшие поиски привели к созданию другой группы теорий, в которых основное внимание уделялось гуморальным факторам мотиваций. Так, голод связывался с возникновением так называемой голодной крови, т. е. крови с существенным отклонением от обычной разницы в концентрации глюкозы в артериальной и венозной крови. Это и рассматривалось в качестве первопричины «голодных» сокращений желудка. Мотивация жажды также оценивалась как следствие изменения осмотического давления плазмы крови или снижения внеклеточной воды в тканях, что может оцениваться специальными рецепторами (осмо- и волюмрецепторы) . Половое влечение ставилось в прямую зависимость от уровня половых гормонов в крови.
Для оценки справедливости гуморальных теорий мотиваций существенный интерес представляют наблюдения за двумя парами сросшихся близнецов-сакропагах, выполненные Т. Т. Алек-
сеевой (1958). У этих близнецов была общая система кровообращения и раздельная центральная нервная система. Верхняя часть пищеварительного тракта у них также была раздельной. Оказалось, что кормление одной из девочек, а значит равномерное распределение питательных веществ в кровеносной системе обоих близнецов, никогда не приводило к насыщению второй девочки. Наоборот, у нее длительное время могла поддерживаться потребность в пище. Эти наблюдения, а также многочисленные наблюдения на животных показали, что гуморальные факторы не могут быть первопричиной возникновения биологических мотиваций.
По-видимому, как возникновение мотиваций, так и их удовлетворение имеет множественный генез, в основе которого лежит нейрогуморальный механизм периферического и центрального уровней.
В самих мозговых структурах обнаружены чувствительные нервные клетки, специализированные на восприятии колебаний в содержании определенных химических веществ, а также физических параметров питающих их жидкостей. Основным аппаратом, содержащим такие центральные рецепторы, является гипоталамус. И. Стелл ар (1954), выдвинув гипоталамическую теорию мотиваций, полагал, что гипоталамус и является вместилищем «центрального мотивационного состояния».
Рассмотрим для примера некоторые гипоталамические механизмы жажды. Во-первых, в латеральных ядрах гипоталамуса обнаружены осморецепторы, детектирующие осмоляльность циркулирующей крови. Во-вторых, введение в латеральный гипоталамус солевого гипертонического раствора вызывает усиление потребления воды сверх физиологических потребностей (похожую реакцию вызывает и электрическое раздражение этих же структур). В-третьих, повреждение латерального гипоталамуса нарушает регуляцию водного баланса, а повреждение переднего гипоталамуса и преоптического ядра полностью устраняет способность к потреблению воды, не лишая, однако, животных воз-
можности принимать пищу. И наконец, в-четвертых, раздражение «питьевой» зоны гипоталамуса провоцирует целый комплекс поведенческих реакций, связанных с поисками воды.
Хорошо известно, что прекращение питья наступает задолго до того, как восстановятся физиологические константы крови. Показано также, что ни растяжение желудка, ни всасывание воды не имеет решающего значения в регуляции потребления воды. По этому поводу высказывается наиболее вероятное предположение — прекращение приема воды наступает в результате обратной связи от рецепторов ротовой полости, с помощью которой отмеряется количество поглощенной воды, и через амигдалу тормозится реакция питья (Н. Мильнер, 1973).
Теперь рассмотрим некоторые теории мотивации голода, которые связывают это состояние опять же со структурами гипоталамуса. Наиболее последовательные поиски ведутся Б. К. Анандом и Д. Р. Бробеком (1951), которые считают, что в латеральном гипоталамусе располагается центр питания, а в медиальном гипоталамусе — центр насыщения. Авторы допускают, что центры питания и насыщения находятся в реципрокных отношениях. Потребление пищи определяется тормозными влияниями латеральной области на медиальную.
С. П. Гроссман (1960—1969) вводил в гипоталамус накормленных крыс биологически активные вещества. Введение в латеральный гипоталамус адренергических веществ (в отличие от холинергических) вызывало усиленное потребление пищи. Введение же холинергических веществ провоцировало питьевые реакции. При этом возникал целый комплекс ранее заученных водо-и пищедобывательных двигательных реакций.
Сегодня является бесспорным положение о том, что
гипоталамические структуры не мoгyт paссматриваться вкачестве единственных аппаратов пищевогоповедения.
Лимбический уровень (включая гипоталамические и ретикулярные механизмы) обеспечивает лишь примитивно
организованные лишенные выраженной целенаправленности типы пищевого поведения, так называемые пищевые автоматизмы. Направленный же поиск пищи, ее оценка и выбор, а также формирование избирательного аппетита требуют участия коры больших полушарий головного мозга. Следовательно, только объединенное участие гипоталамуса, лимбических образований и коры головного мозга формирует целостное пищевое поведение животных.
При этом исключительная роль гипоталамуса не вызывает никаких сомнений:
Этот отдел мозга как комплексный детектор физиологических дисбалансов (температуры, питательных веществ, воды, гормонов, минеральных солей) играет доминирующую роль в отношении путей использования ресурсов организма.
Практически ни одну функцию организма нельзя себе представить вне прямой или косвенной зависимости от гипоталамуса.
Работами К. В. Судакова (1971, 1986) было показано, что каждое мотива ционное возбуждение строится на основе специфических химических механизмов, активирующих влияния подкорковых мотивационных центров на кору головного мозга. Введением центральных адренолитиков (аминазина) удается избирательно заблокировать активацию коры при болевом раздражении. Активация же коры головного мозга при пищевом возбуждении голодных животных избирательно блокируется холино-литическими веществами.
Наряду с вышесказанным существует и крайняя точка зрения П. Мак-Лина (1970) о жесткой привязке мотиваций к аппаратам лимбической системы. Он считает, что в гипоталамусе локализованы эмоционально окрашенные поведенческие реакции, связанные с рефлексами избегания, нападения, поиска, добывания пищи, воды и пр. В амигдале сосредоточены мотивации голода, жажды, защиты от внешних вредных влияний. Перегородка связана с половым поведением. Противоположной позиции, которая кажется нам более справедливой, придерживается Р. Л. Аисааксон (1978), который отмечает, что не существует
одиночных анатомических образований для одиночных поведенческих функций, как не существует единых поведенческих функций, которые могли бы быть приурочены к анатомическим образованиям таким образом, чтобы эти функции оказывались полезными в любых обстоятельствах и при всех внешних условиях.
Доминирующее мотивационное возбуждение.В каждый момент времени доминирует та мотивация, в основе которой лежит наиболее важная биологическая потребность. Сила потребности, т. е. величины отклонения физиологических констант или концентрации соответствующих гормональных агентов, получает свое отражение в величине мотива ционного возбуждения структур лимбической системы и определяет его доминантный характер. Консервативный характер доминанты заключается в ее инертности, устойчивости, длительности. В этом ее большой биологический смысл для организма, который стремится к удовлетворению биологической потребности в случайно и постоянно меняющейся внешней среде. В физиологическом смысле такое состояние доминанты характеризуются определенным уровнем возбудимости центральных структур, обеспечивающей их высокую отзывчивость и «впечатлительность» к разнообразным воздействиям. Но однажды проявленная доминирующая мотивация возобновляется и живет даже в порядке чисто нервно-рефлекторного фактора. Поэтому наряду с консервативным началом в доминирующей мотивации как векторе поведения содержится и динамическое начало, поскольку из множества новых подкрепляющих впечатлений происходит непрерывный подбор «пригодного», нужного, имеющего непосредственную связь.
Первичный очаг возбуждения при формировании доминирующих биологических мотиваций (голод, жажда, секс) возникает в гипоталамических образованиях мозга (Р. А. Павлыгина, 1956). Эти первичные очаги, в свою очередь, ведут к появлению вторичных очагов в других отделах мозга, где

Рис. 21. Схема соотношения первичных и вторичных очагов при формировании доминирующей констелляции моягопых образований (по Л. С. Батуеву, 1986)
имеются предпосылки для продолжительного удержания следовых процессов. Кора головного мозга с ее непрерывной и разномодальной активацией располагает всеми предпосылками для создания в ней вторичных очагов стационарного возбуждения. Принципиальным отличием таких очагов от первичных является исходная причина их возникновения. Так, если первичный очаг в гипоталамусе возникает и поддерживается в первую очередь за счет гуморальных факторов, то вторичные очаги в коре больших полушарий являются проекционными по своему происхождению и поддерживаются за счет широкой конвергенции к ним множества суммирующихся здесь сенсорных влияний.
Рассмотрим несколько конкретных примеров. Содержание животных на длительной солевой диете, формирующее у них мотивацию жажды, создает в латеральном гипоталамусе очаг повышенной возбудимости к вкусовым и внсцерохи ми чески м раздражениям
(А. С. Батуев, Б. Г. Гафуров, 1988). Снимается же подобный очаг введением в организм дополнительных количеств жидкости.
Ф Итак, мотивационное возбуждение, обусловленное преобладающей потребностью организма, формирует в гипоталамусе первичный очаг повышенной возбудимости, обладающий свойствами доминантного очага. Благодаря этому доминантному очагу быстро образуются условно-рефлекторные связи с теми или иными фа.кторами окружающей среды (вторичные очаги), которые по своей функциональной значимости могут быть сравнимы с эндогенными механизмами самого мотива-ционного состояния (рис. 21).
Нейрофизиология мотиваций.К. В. Судаков (1986) сформулировал основные положения о принципах нейрофи-зиологического обеспечения доминирующих мотиваций: 1. Любая биологическая мотивация обусловлена соответствующей метаболической потребностью и носит системный характер.
2. Потребность трансформируется ней-рогуморальным путем в возбуждение гипоталамических центров, которые активируют другие структуры мозга, в том числе и кору больших полушарий.
3. Корковые и лимбические структуры мозга оказывают специфические для каждой мотивации нисходящие возбуждающие и тормозные влияния на гипоталамические мотивационные центры. 4. Каждое мотивационное возбуждение представляет собой специфическую клеточную и молекулярную интеграцию корково-подкорковых структур. Избирательное возбуждение одних структурных образований сочетается с избирательным торможением других.
Мотивационные возбуждения существенно меняют конвергентные и дискриминационные способности нейронов разных отделов мозга. Например, раздражение пищевого центра латерального гипоталамуса приводило к тому, что нейроны сенсомоторной коры, ранее не отвечавшие на световые,
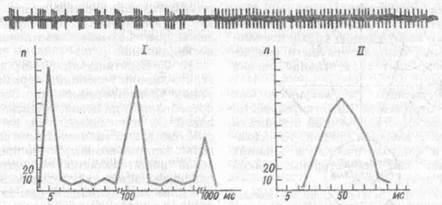
Рис. 22. Типичный характер нейронной активности у голодного (/) и накормленного кролика (//) (по К. В. Судакову, 1986):
под нейронограммой — гистограммы распределения межимнульсных интервалов
звуковые и гуморальные раздражения, приобрели способность реагировать на них. С другой стороны, нейроны коры, не отвечавшие ранее на введение кроликам морковного сока, начинали реагировать на эти раздражения после стимуляции центра «голода» латерального гипоталамуса. Создается впечатление, что мотивационное возбуждение «настраивает» нейроны различных областей мозга на подкрепляющие воздействия.
Мотивационное возбуждение находит свое выражение в специфическом распределении межимпульсных интервалов и образовании характерной пачечной активности (рис. 22). Эта активность усиливается при возрастании потребности, при возникновении препятствий к удовлетворению доминирующей мотивации. Достижение же цели поведенческого акта сменяется в нейронограмме регулярной активностью.
Возбуждения, поступающие в мозг при подкреплении, адресуются к различным нейронам и оставляют «след» в их деятельности, формируя сложную нейродинамическую констелляцию. С ней и происходит сравнение параметров достигнутых результатов.
В. Г. Зилов (1978) показал, что в формировании различных биологи-
ческих мотивации участвуют одни и те же нейромедиаторы, однако в различных комбинациях в разных структурах мозга, что свидетельствует о специфической нейрохимической интеграции конкретного мотивапионного возбуждения. Последнее проявляется также и в общей чувствительности отдельных нейронов к нейромедиаторам и оли-гопептидам (К. В. Судаков, 1986). Способность ряда пептидов (ангиотензин И, бета-липопротеин, пентогастрин и др.) при внутрижелудочковом введении избирательно продуцировать или подавлять определенные биологические мотивации открывает путь к изучению молекулярной интеграции мотива-ционного возбуждения.
Дата добавления: 2015-08-04; просмотров: 2563;