Естественный отбор благоприятствует группировке тех генов, которые способствуют проявлению полезного поведенческого признака.
Эффективным путем для изучения эволюции поведения является вышеописанный прием экспериментального анализа наследования КФД. ЛАетод получения межвидовых гибридов оказывается в этом плане весьма убедительным.
Нервно-психические заболевания генетической природы.К настоящему времени известно более 80 заболев'а-ний, связанных с определенными генетическими дефектами. И хотя очевидно, что наследственная информация влияет на развитие психопатологических симптомов, конкретные механизмы этого влияния в большинстве случаев остаются неизвестными.
ф Определенные виды расстройств высшей нервной деятельности связывают с тремя типами генетических аномалий. 1. Аномалии, связанные с рецессивными генами, возникают в результате дефицита определенного фермента в печени. Типичным примером такого рода заболевания служит фенил-кетонурия, которая в поведении проявляется в форме задержки умственного развития, а иногда сопровождается эпилептическими припадками и психо-
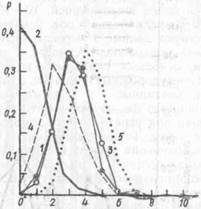
Рис. 17. Вероятность совершения крысами определенного числа ошибок при поиске пищи в лучах радиального лабиринта (по А. С. Бятуеву и др., 1983);
По оси ординат — вероятность; по оси абсцисс — число ошибок; / — средние величины у «необученных» (1 — 10-й опыты) и 2- у обученных (47 --56-й опыты) крыс линии Вистср; 3 — у «необученных» и 4 — у обученных крыс линии Крут и некого -Молодкиной; 5 — математическая мидель случайных посещений
зами. Данное заболевание обусловлено изменением генетического кода в одном участке хромосомы. 2. Аномалии, связанные с доминантными генами, в заболевании проявляются при особых условиях или стрессах. Ярким примером может служить хорея Геттингтона, которая начинается обычно в возрасте 30—40 лет (дегенерация коры и базальных ганглиев) и проявляется в непроизвольных движениях, неправильной подергивающейся походке, характерных гримасах, замедленной, невнятной речи, ухудшении памяти, повышенной раздражительности, депрессии, слабоумии.
Сходны с этой болезнью и такие наследственные заболевания нервной системы, как болезнь Альцгеймера и болезнь Паркинсона (дрожательный паралич). Допускают, что такую же природу имеют шизофрения (грубое расстройство интеллектуальной деятельности) и маниакально-депрессивные психозы. Высокая наследственная предрасположенность к шизофрении не вызывает сомнения. Риск заболевания
шизофренией у детей, где один из родителей болен шизофренией, 10—15 и 35—40 %, если больны оба родителя. Существуют данные о высокой наследственной предрасположенности к эпилепсии, основным симптомом которой являются судорожные припадки. 3. Аномалии, связанные с хромосомными аберрациями, заключающимися в добавлении или утрате целой хромосомы или ее части. Типичным примером такого заболевания служит болезнь Дауна, выражающаяся в задержке умственного и психического развития. Больной имеет 47 хромосом вместо 46, свойственных нормальному человеку. Наличие лишней хромосомы обусловливает синтез избыточного количества фермента, необходимого для построения белков головного мозга.
Группу заболеваний с ранним нарушением интеллекта объединяют под термином олигофрения. При этом хромосомные мутации могут выражаться в изменении числа хромосом, их структуры или краткого изменения гаплоид-ного набора. И всегда они сопровождаются тяжелыми формами умственной отсталости в сочетании с различными пороками развития.
Л. Г. Романова и Л. Г. Калмыкова (1981) выдвинули гипотезу о генетической природе психических болезней. Они полагают, что доминантные мутации должны приводить к поражению или дисфункции строго ограниченных структур мозга. Приуроченность данного психического дефекта к определенным жестким мозговым системам дает возможность понять не только мозговые механизмы психических расстройств, но и определенные аспекты нормального поведения.
Генетика свойств нервных процессов.Первым, кто обратил внимание на необходимость изучить генетическую природу свойств основных нервных процессов (их силы, уравновешенности и подвижности), был И. П. Павлов в связи с разработкой им физиологической базы для определения типов нервной системы животных.
В дальнейшем работами отечественных ученых (Л. В. Крушинский, 1946;
В. К. Красусский, 1963) была доказана генетическая детерминированность силы возбудительного процесса, а В. К. Федоров (1971) показал зависимость от генотипа материнского организма такого свойства, как подвижность нервных процессов.
М. Е. Лобашов обратил внимание на наследование такого фундаментального свойства нервной системы, как возбудимость, которая определяет действие генов, детерминирующих отдельные поведенческие акты.
Ученики М. Е. Лобашова создали стройную систему взглядов на нейрохимическую и молекулярную детерминацию поведения (Н. Г. Лопатина, В. В. Пономаренко, 1987).
Исследования позволили вычленить эффект отдельного гена, вносящего основной вклад в формирование порога нервно-мышечной возбудимости, а также установить его идентичность с определенным локусом хромосомы. Более того, эти же гены участвуют в наследственной детерминации пороговых характеристик нервной системы в целом. У видов, пород и рас животных, имеющих высокую нервно-мышечную возбудимость, наблюдали и более высокую пищевую возбудимость и более высокие показатели силы возбуждения. Основой такой общности генетических механизмов могут быть функциональные или структурные особенности клеточных мембран и связанных с ними ферментов, определяющих характер протекания внутриклеточных процессов (В. В. Пономаренко, 1970).
Генетическая детерминация особенностей обучения.Выведены линии крыс, различающихся по обучаемости в сложном лабиринте: способные крысы ориентировались лучше по пространственным ориентирам, а неспособные — по зрительным. Большое значение при этом имеет характер мотивации: способные крысы сильнее мотивируются голодом, а неспособные — в защитно-оборонительных ситуациях.
Вклад каждой из скрещиваемых линий в свойства обучаемости конкретного поколения и особи неодинаков. М. П. Садовникова-Кольцова (1928)
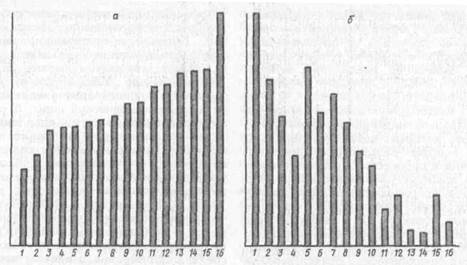
Рис. 18. Корреляции между порогом Возбудимости нервно-мышечного аппарата и способностью к образованию условного рефлекса активного избегания у мышей 16 ннбредных линий (по Л. С. Дмитриеву, 1083);
А— порог возбудимости при раздражении икроножний мышцы одиночными электрическими импульсамидлительностью 2 мс; б — количество условных рефлексов активного избеганияr %; по горизонтали — линии мышей
предположила наличие у крыс трех групп генов, детерминирующих способность к обучению: гены двигательной активности, гены эмоции страха и гены ориентировочно-исследовательского инстинкта. Однако корреляция этих свойств со способностью к обучению не всегда проявляется, и в настоящее время рассматривают эти комбинации признаков как носящие фенотипический характер.
Наследственную основу различий в обучении М. Е. Лобашов предполагал искать в генетически детерминированных особенностях безусловных рефлексов и прежде всего в уровне пищевого рефлекса как основного модулятора функциональной активности нервной системы. Показано наличие генетических корреляций между уровнем функциональной активности нервной системы и способностью к выработке условных рефлексов (В. В. Понома-ренко).
Ю. С. Дмитриевым (1981) было установлено, что порог возбудимости и скорость образования оборонительных условных рефлексов проявляют сходный характер наследования (рис. 18).
Вероятно, в основе лежит множественный эффект какой-то общей группы генов. В число контролируемых им признаков поведения следует отнести ориентировочно-исследовательскую активность, общую двигательную активность и уровень эмоционального статуса. Последние могут участвовать в наследственной обусловленности условно-рефлекторных форм поведения. Ген, контролирующий вышеназванные признаки, идентичен гену, определяющему содержание биологически активных соединений в плазме крови, серотонина и норадрсналииа в тканях гипоталамуса (А. Оливерио и др., 1973; Ю. С. Дмитриев, 1981).
ф Таким образом, множественное влияние одного и того же гена может заключаться в контроле порога возбудимости нервной системы, содержания нейроактивных соединений и способности к обучению (образованию оборонительных условных рефлексов).
Генотип и анатомические особенности мозга.Благодаря трансплантации появилась возможность экспериментального изменения массы мозга. Пересадка африканской рыбке-теляпии
среднего мозга от донора увеличила общую массу мозга реципиента и одновременно улучшила ее интеллектуальные способности: рыбка успешно выполняла задачи по различению и переделке сигнального значения раздражителей, что до операции делала с трудом. К настоящему времени получено значительное число убедительных аргументов, которые позволили приписывать особую роль структурам гип-покампа в процессах обучения и памяти. В процессе начальных этапов обучения в нейронах гиппокампа наблюдается более интенсивное включение меченых белков (Р. Хиден, 1972). Ухудшение способности к обучению связывают, соответственно с нарушением синтеза специфических белков нервной ткани (типа S-100). Если в наследственной обусловленности структурных признаков и способности к обучению участвуют общие гены, то
генетическая изменчивость одного из признаков повлечет за собой изменения и в уровне проявления второго.
Оказалось, что изменчивость синапсов на клетках гиппокампа значима для наследственно обусловленного уровня активации этих нейронов. Причем число окончаний мшистых волокон на денд-ритах пирамидных клеток находится в определенной связи со способностью мышей к образованию оборонительных условных рефлексов. Между числом терминален на базальных дендритах и уровнем осуществления условных рефлексов существует высокая отрицательная корреляция. Высокий уровень условно-рефлекторной деятельности коррелирует с многочисленностью синапсов на апикальных дендритах пирамид гиппокампа. Последнее связывают с тем, что создаются лучшие условия для проявления состояния длительной активации гиппокампа, что обеспечивает его участие в процессах обучения и запоминания.
Структурные изменения распространяются и на другие отделы мозга: у крыс с высоким уровнем условно-рефлекторной деятельности обнаружена большая ширина сенсомоторной области коры, большие размеры зубча-
той фасции, мозолистого тела с большим числом миелинизированных волокон. Генетически детерминированные структурные особенности захватывают и лимбическую систему мозга (Н. И. Дмитриева и др., 1983, 1985): а) у хорошо обучающихся крыс увеличивается ширина лимбической коры; б) увеличивается размер клеток ядер гипоталамуса и амигдалы; в) увеличивается число глиальных клеток свода по сравнению с животными с низким уровнем возбудимости и скорости образования условных рефлексов.
Л. В. Крушинский (1977) считает, что избыточное количество нейронов мозга — необходимое условия для полноценного восприятия окружающей среды и формирования адекватных поведенческих реакций.
Генотип и нейрохимия мозга.Наследование свойств обучаемости по материнской линии позволяет обратить внимание на значение не только ядерного аппарата нервных клеток, но и всего цитоплазм этического содержимого. Реализация генетической информации, закодированной в молекуле ДНК в ядре нервной клетки, осуществляется при непосредственном участии химических факторов самой цитоплазмы клетки. Помимо широко известных первичных химических посредников-нейромедиаторов, с помощью которых информация передается к нервной клетке и активирует ее в соответствии с присущей ей собственной генетической программой, в настоящее время в самостоятельную категорию метаболических факторов выделены вторичные посредники (мессенджеры). В первую очередь к ним относят циклический аденазинмо-нофосфат(цАМФ), выполняющий функцию универсального клеточного регулятора.
Нейромедиаторы и пептидные гормоны выступают по отношению к клетке в качестве сигналов. Эти сигналы воздействуют на мембрану клетки и способствуют образованию из АТФ вторичных посредников — цАМФ, который стимулирует более чем 200-кратное усиление воздействия на клетку внешнего для нее сигнала. цАМФ активи-
рует соответствующие ферменты (про-теинкиназы), обеспечивающие образование определенных белков. Эти рецепторные белки встраиваются в по-стсинаптическую мембрану клетки и способствуют высокой эффективности воздействия определенного сигнала на клетку.
Ионы кальция также относят к категории вторичных посредников, от которых зависят как пре-, так и постсина-птические процессы клетки и формирование ее электрической активности. Вслед за открытием рецептора кальция — белка кальмодулина было установлено, что он регулирует синтез и распад цАМФ.
Особое внимание обращают на сте-роидные гормоны, которые реализуют свои эффекты, минуя систему вторичных посредников. В отличие от пептид-ных гормонов стероидные гормоны уже имеют собственные возможности проникновения в нервную клетку, где они связываются непосредственно с се ядром. Три основных фактора регуляции: вторичные посредники, ионы кальция и стероидные гормоны — взаимодо-полняют друг друга.
В. В. Пономаренко (1970) предложила гипотезу нейроэндокринной регуляции процесса реализации генетической информации, согласно которой вторичные посредники и стероидные гормоны представляют собой интеграцию функций нервной и эндокринной систем на молекулярном уровне. Возможность регуляции активности генов в нервной системе возрастает в эволюции за счет увеличения как нейрональных, так и гормональных звеньев этой регуляции, а также онтогенетического влияния на эти звенья, включая весь приобретенный индивидуальный опыт.
Гетерохрония реализации генетической программы.Признаки и свойства мозга, детерминируемые генетической программой, образуют так называемую консервативную наследственность вида. Она может отражаться в структурных или функциональных признаках, в константной последовательности этапов развития поведения. Эти признаки инвариантны для целого вида и возникли
иод действием стабилизирующего отбора. Динамика онтогенеза является удобной моделью для изучения закономерностей развертывания генетической программы. Например, изучение созревания восходящих возбуждений в кору мозга от ретикулярной формации, от зрительной системы и пр. позволяет представить ход развития врожденных матриц будущих поведенческих актов, формирующихся в процессе обучения (Ф. А. Ата-Мурадова, 1983). Особенность онтогенетического метода заключается во вмешательстве в гетерохронную активацию генома (гл. 3, §10).
Биогенетическая последовательность стадий развития физиологических систем отражает стадийное действие генных факторов различного эволюционного возраста. Активность генов каждой стадии развития, формирующей соответствующий фенотипический признак, создает необходимый генетический фон и включает новые системы генов следующей стадии. Например, созревающие первыми структуры восходящих корковых возбуждений прокладывают путь для других компонентов поздно созревающих структур.
П. К. Анохин (1968) подчеркивал опережающее активирование специфической генетической системы, определяющей стадийность развития, и раннее включение нейрональных функций как системного процесса. Так, синхронизация активности в популяциях нейронов, объединяющихся в рано созревающие ансамбли, отражает системный характер активации генетической программы. Установлена первичность созревания неспецифических восходящих в кору возбуждений. Вначале созревает более древняя холинергическая форма проведения, которая обусловливает первичную реакцию гиперсинхронизации. Затем добавляется адренсргиче-ский механизм, заменяющий гиперсинхронизацию характерной для взрослого десинхронизацией (Ф. А. Ата-Мурадова, 1983). При этом подчеркивается жесткость генетической запрограммированности данных феноменов, что свидетельствует об известной инвариантно-
ста соответствующих генных интеграции и об их значительном, историческом возрасте.
Одной из иллюстраций такого положения могут служить эксперименты с пересадкой (трансплантацией) мозговой ткани у амфибий. Пересадка закладки головного мозга между зародышами разных видов бесхвостых амфибий, значительно различающихся по поведению, показывает, что поведение в основном определяется трансплантированным мозгом (Л. В. Полежаев, 1983). Например, прошедшие метомор-фоз остромордые лягушки с пересаженным им мозгом жабы-чесночницы демонстрируют поведение жабы-чесночницы: ползают на брюхе, выкапывают в земле ямки и зарываются в них.
*
Дата добавления: 2015-08-04; просмотров: 1493;