Структура и типы мышечных волокон.
Структура поперечно-полосатой соматической мышечной ткани позвоночных животных
Анатомическая структура поперечно-полосатой мышечной ткани
Поперечно-полосатая соматическая мышечная ткань состоит из симпластических мышечных волокон, образующих мышцу.
Мышца, как анатомическая единица мышечной системы позвоночных животных и человека, содержит различное количество мышечных волокон разного типа (см: лекцию № 1), объединенных в пучки и окружена системой оболочек.

Рис. 2.1.1 Общая организация соматической мышцы позвоночных. Показаны уровни анатомической и гистологической организации мышечной ткани.
Вся мышца окружена мощной соединительно-тканной оболочкой, (эпимизием), содержащей многочисленные кровеносные и лимфатические сосуды, нервы, жировые элементы, мощные пучки коллагеновых и эластических волокон, которые на концах мышцы образуют сухожилия (tendon) , прикрепляющие мышцу к кости.
Отдельные группы мышечных волокон внутри мышцы окружены более тонкой соединительно-тканной оболочкой – перимизием,а отдельные мышечные волокна отделены друг от друга тонкими прослойками внутренней оболочки – эндомизия.
Крупные мышцы имеют более толстую наружную соединительно-тканную оболочку, называемую фасцией (рис. 2.3).Онадвуслойнаяи состоит из собственно эпимизия, прилегающего непосредственно к мышце и наружного листка фасции, контактирующего с окружающими органами и тканями. Между этими листками есть узкое пространство, заполненное небольшим количеством тканевой жидкости и предназначенной для уменьшения трения при сокращении мышцы.
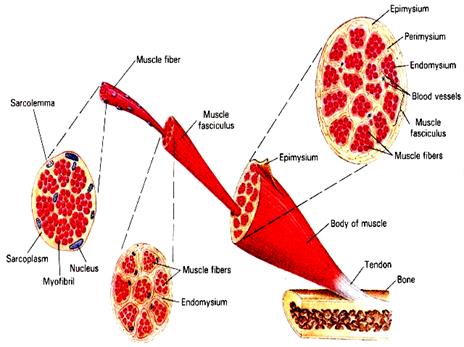
Рис. 2.1.2. Микроанатомическое строение мышцы и ее оболочек.
Соединительно-тканная оболочка мышцы и отдельных мышечных волокон имеет достаточно сложную структуру.
Артерии и капилляры проникают в мышечную ткань через эпимизий, далее в перимизий, далее вдоль мышечного волокна в составе эндомизия. На одно мышечное волокно приходится 2-4 капилляра. Они образуют богатую сеть анастомозов. Там-же располагаются вены и лимфатические сосуды и капилляры.
Сарколемма мышечного волокна состоит из собственно плазматической мембраны волокна, базальной мембраны и тонкого слоя эндомизия. Базальная мембрана представлена аморфным веществом из протеогликанов и гликопротеинов (фибронектин, ламилин, энтактин и др) и тонких коллагеновых фибрилл IV типа, диаметром 4-5 нм.
Снаружи от базальной мембраны располагается система фибриллярных структур, образующих своеобразный каркас вокруг волокна. Этот каркас состоит из фибриллярных белков (Рис.2.1.3 ).Этот каркас выполняет достачно важные функции в мышце. Во-первых он обеспечивает деформацию и одновременно сохраняют целостность волокна в продольном и поперечном направлениях при сокращении мышцы. Во-вторых, он, наряду с системой внутриклеточных цитоскелетных структур, обеспечивает упругость волокна, препятствует его искривлению, гасит ударные нагрузки и вибрации. Наконец – этот своеобразный каркас для проходящих здесь капилляров, которые при сокращении волокон сильно деформируются
Рис 2.1.4 ).
Выше этого каркаса распологается плотная сеть тонких коллагеновых (IV типа) волокон диаметром 200-250нм, с одной стороны проникающие в базальную мембрану, с другой переплетающиеся с волокнами эндомизия. Сам эндомизий состоит из сети микрофиламентов в 7-10 нм, тонких коллагеновых фибрилл в 50-60 нм и толстых коллагеновых фибрилл (коллаген I,III,IV,V типов) различного диаметра и эластических волокон, не контактирующих с базальной мембраной, а переплетающиеся с волокнами перимизия.
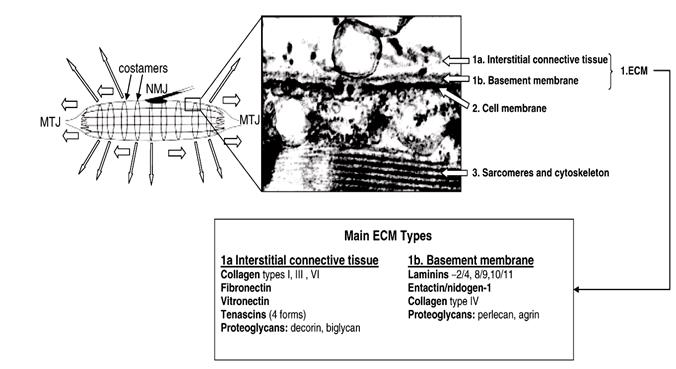
Рис. 2.1.3. Система цитоскелетного каркаса вокруг мышечных волокон
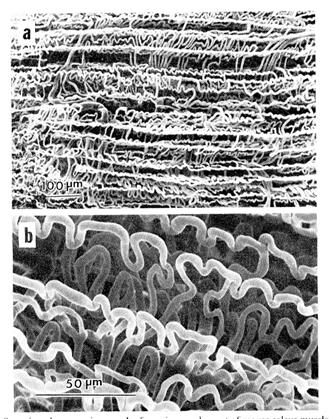
Рис. 2.1.4. Капиллярная сеть мышечного волокна, находящегося в сокращенном состоянии ( по: H.Ishikawa, Cell and muscle motility, 1983. vol.4).
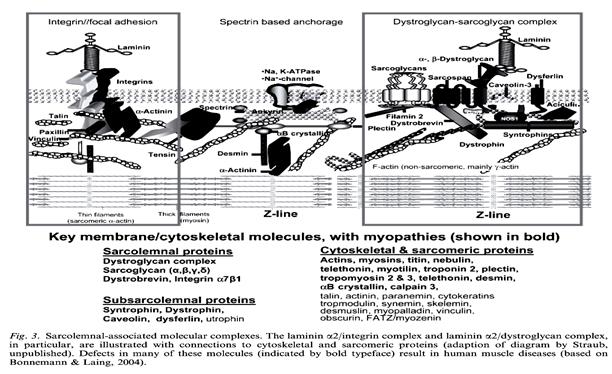
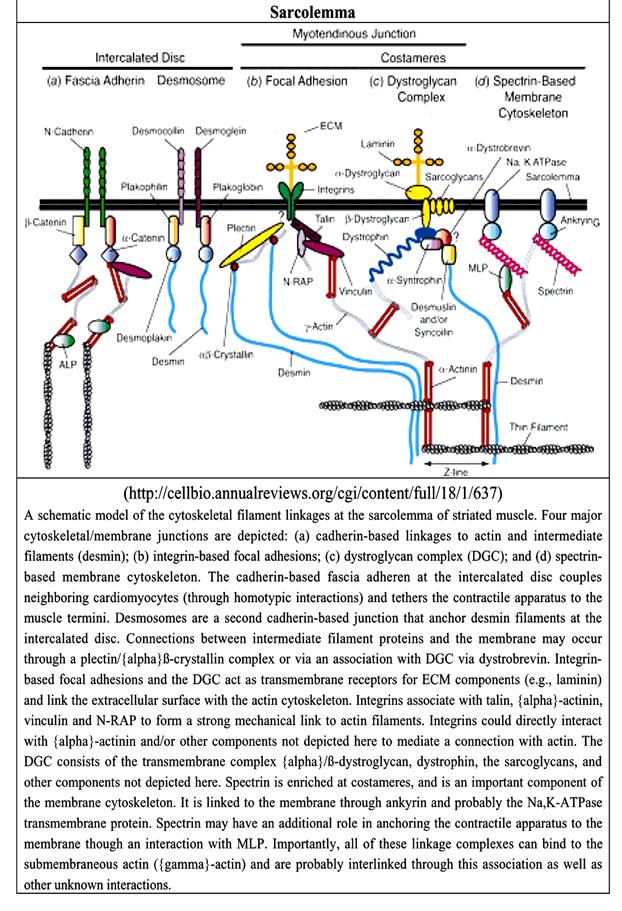
Рис. 2.1.5. Схема строения и состав сарколеммы мышечного волокна
(по: E.Niederhoffer, Muscle diochemistry,2004, SI University Press)
Структура и типы мышечных волокон.
Мышечные волокна соматической (скелетной) мышцы позвоночных имеют симпластическое строение и включают в свой состав несколько десятков ядер слившихся в процессе формирования мышечного волокна миоцитов.
Длина мышечных волокон сильно варьирует в зависимости от мышцы, в состав которой они входят и, в среднем, составляет от сотен микрон до 10-15 см с диаметром в несколько десятков или сотен микрон.
В составе мышцы, помимо белых (IIB), красных (I) и промежуточных (II A) мышечных волокон, которые получили наименование «экстрафузальных» и о которых рассказывалось выше, есть т.н. «интрафузальные» мышечные волокна.

Рис. 2.2.1. Микрофотография отдельного соматического мышечного волокна (вверху) и пучка волокон (внизу). Видна поперечная исчерченность и многочисленные ядра волокна и ядра миосателлитов, а также ядра фибробластов, образующих эндомизий.
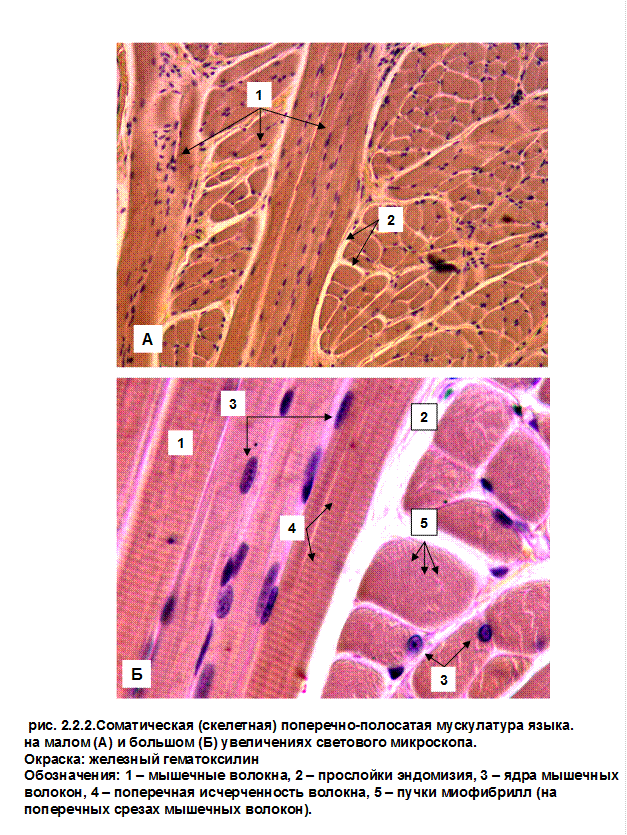
Интрафузальные мышечные волокна входят в состав мышцы и предназначены для обеспечения нервной «настройки» мышцы. Они имеют небольшой диаметр и малочисленны, не создавая выраженного мышечного усилия при сокращении. Располагаясь в толще мышцы они образуют пучки волокон – «мышечное веретено», окруженное собственной оболочкой.
Число волокон в интрафузальном веретене у разных животных разное:
- у рептилий -1
- у лягушек – 3-12,
- у крыс – 4,
- у кошек – 2-13,
- у человека – 3-7.
Мышечные веретена представляют собой пучок волокон, окруженных отдельной внутренней соединительно-тканной капсулой. Волокна двух типов : волокна с скоплениями ядер в центральной части волокна – ядерно-сумковые волокна (медленные) и волокна с ядрами, рассеянными по всей длине волокна – ядерно-цепочечные (быстрые).Кроме внутренней капсулы, район мышечного веретена окружен наружной капсулой,. в пределах которой также находятся кровеносные сосуды и пучки периферических нервов.
У человека в веретене волокон я ядерной сумкой – 1-3, с ядерной цепочкой – 3-7. Ядерно-сумковые волокна выходят за пределы капсулы и прикрепляются к эндомизию, тогда как ядерно-цепочечные волокна короткие и не выходят за пределы капсулы.
Мышечные веретена реагируют на скорость сокращения мышцы и ее растяжение, имея в своем составе многочисленные чувствительные нервные окончания. Иннервация осуществляется двумя типами нейронов:
- первичными афферентными нейронами (группа 1а). Их волокна относятся к высокоскоростным афферентам (120 м/сек) и образуют спиралевидные окончания в центре интрафузальных волокон. Они реагируют, в основном, на скорость растяжения мышцы при действии нагрузки.
- вторичными афферентными нейронами (группа 2), рецепторные окончания которых располагаются на полюсах ЯЦ волокон. Они реагируют на постоянное напряжение мышцы и практически не реагируют на растяжение мышцы.
Эфферентная (двигательная) иннервация интрафузальных волокон осуществляется γ – мотонейронами, образующими тонкие (2-4 мкм) двигательные нервные волокна ( А- типа), называемые «фузимоторными».
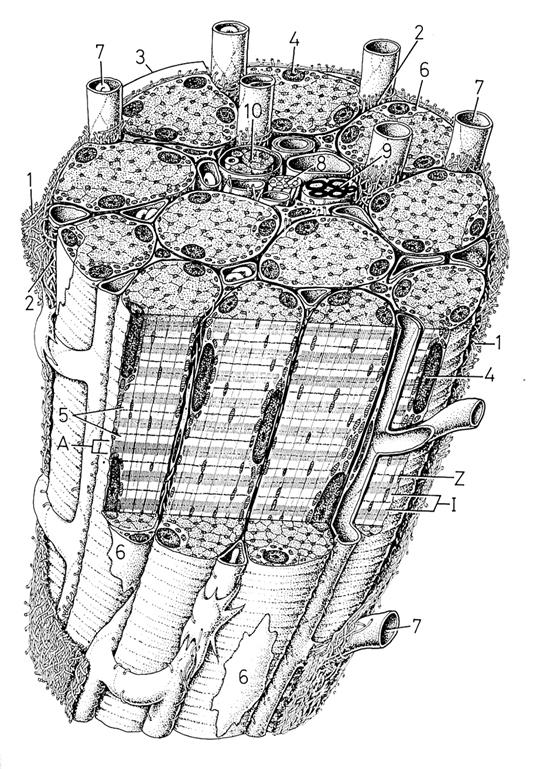
Рис. 2.2.3 Строение мышцы. В центре виден центральный пучок из нервов, кровеносных сосудов и интафузальных волокон (мышечное веретено - 10) (по R.Krstic, 2001)

Рис. 2.2.4. Гистология участка мышцы позвоночных, включая экстрафузальные и интрафузальные мышечные волокна. Участок «мышечного веретена», содержащий интрафузальные мышечные волокна обведен кругом.

Рис. 2.2.5. Схема строения «мышечного веретена» и система его иннервации (по: Гистология, 2001)
2.3. Саркомер – как основная морфо-функциональная единица мышечного волокна.
Саркомер является структурной единицей миофибрилл поперечно-полосатого мышечного волокна соматической (скелетной) мышцы позвоночных , а также ряда мышц беспозвоночных животных и миофибрилл кардиомиоцитов. (Рис. 2.3.1

Рис. 2.3.1. Микрофотография (скан.эл.микр.) и фото саркомера скелетной мышцы.
Длина саркомера составляет 2,5 – 3 мкм, диаметр – 1-2 мкм и выглядит как система чередующихся из дисков. Формула саркомера: Z+½ I + A +½ I + Z.
Он представляет собой систему тонких (thin, actin) актиновых и толстых (thick, myosin) филаментов
А - диск –зона локализации толстых миозиновых филаментов. Включает в свой состав несколько подзон:
М – линия (зона крепления миозиновых филаментов в центре саркомера)
Н – подзона (диск) -участок расположения основной массы миозиновых протофибрилл. В пределах этой зоны, исходя из положения одного из важнейших белков саркомера – титина,выделяют ряд участков: С – участок,где нет перекрытия с актиновыми нитямии D – участок,где миозиновые фибриллы истончаются и есть перекрытие с концами актиновых филаменто
I - подзона (диск) –участок расположения актиновых фибрилл вне зоны перекрытия с миозиновыми фибриллами. В каждом саркомере располагается с двух сторон ½ I диска.
Z - линия (диск) –граница соседних саркомеров.
Современные исследования структуры и химического состава саркомеров показали, что они имеют очень сложную пространственную организацию, а в его состав, помимо основных сократимых белков актина и миозина входит более двух десятков вспомогательных белков, принимающих участие в функционировании саркомера и мышцы в целом.
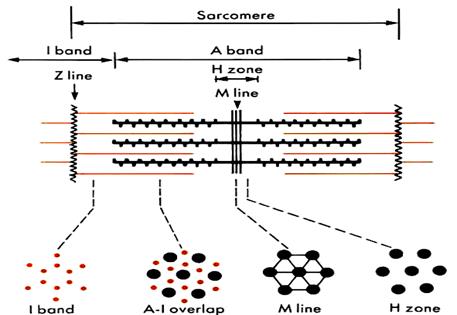
Рис. 2.3.2. Традиционная схема саркомера и его деления на диски

Актиновые и миозиновые протофибриллы образуют сложную упорядоченную сеть. У позвоночных животных встречается два типа «актин-миозиновой решетки», хотя соотношение актин/миозин всегда одно 3:1.
- «суперупорядоченная решетка», характерная для высших позвоночных (млекопитающих и птиц).
- «простая решетка»,характерная для саркомеров мышц низших позвоночных (рыб и амфибий)

Рис.2.3.3. Взаиморасположение актиновых и миозиновых фибрилл
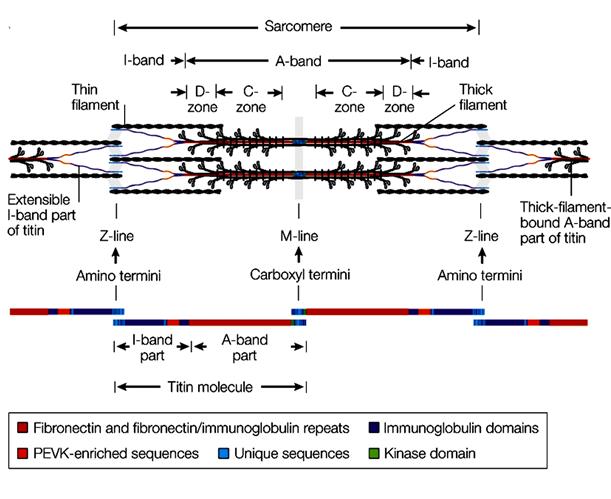
Рис. 2.3.4. Современная схема организации саркомера и расположения в нем белка титина ( по: L.Tskhovebova, J.Trinick, 2003; Physiol.Rev, 2005)
Саркоплазматический ретикулум – представляет собой специализированную сеть гладкого эндоплазматического ретикулума. (L – система). Канальцы СР идут вдоль миофибриллы, но канальцы соседних саркомеров изолированы друг от друга, тогда как СР соседних миофибрилл имеют систему соединительных трубочек. Продольные канальцы СР охватывают миофибриллу со всех сторон, образуя т.н.»воротничок».
По ходу мышечного волокна плазмалемма образует систему поперечных впячиваний – Т- каналов (Т-система). В районах Т-каналов канальцы саркоплазматического ретикулума сливаются, образуя т.н. «терминальные цистерны».Т-каналы могут находится как на уровне Z- дисков (рыбы, амфибии), так и по длине саркомера на уровне А и I –диска (круглоротые, рептилии, птицы и млекопитающие).
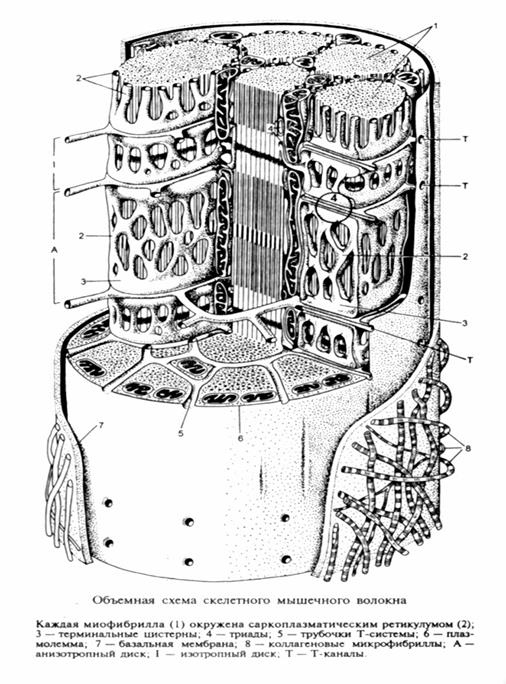

Рис. 2.3.5. Молекулярная структура саркомера (по:E.Niederhoffer, Muscle biochemistry, 2004).
Таким образом каждая трубочка Т-системы с двух сторон контактирует с терминальными цистернами L – системы в пределах каждого саркомера, образуя т.н. «триады».Система L – и Т- каналов значительно различается в тонических и фазных волокнах.
В фазных волокнах, как отмечалось выше, триады располагаются в каждом саркомере не только на уровне Z – дисков, тогда как в тонических мышцах, триады всегда располагаются только на уровне Z – дисков (Кроленко, 1975).
Митохондрии– мышечного волокна очень хорошо развиты, что связано со значительными энергетическими затратами при движении. В связи с этим, в мышцах встречается особый тип митохондрий – митохондриальный ретикулум.
Он представляет собой гигантские разветвленные митохондрии, оплетающие миофибриллы. Между этими пластами ретикулярных митохондрий расположены своеобразные мостики из т.н.нитчатых митохондрий. Тем самым образуется трехмерная митохондриальная сеть, части которой соединены специальными межмитохондриальными контактами (ММК). Через эти контакты возможна связь между отдельными, хотя и гигантскими ретикулярными и нитчатыми митохондриями, объединяя их в единую энергетическую систему мышечного волокна.
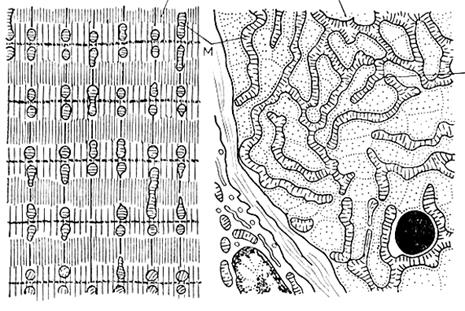
Рис. 2.3.6. Расположение митохондрий в мышце млекопитающих
А – продольный срез, б – на уровне Z – диска (Ченцов, 1995).
Помимо гигантских ретикулярных митохондрий, в мышечных волокнах встречаются митохондрии традиционного типа – отдельные, мелкие митохондрию Особенно много их в околоядерной области.
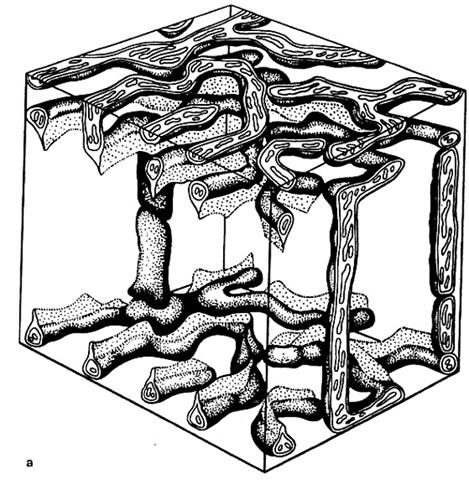
Рис. 2.3.7. Объемная реконструкция участка митохондриального ретикулума в мышечном волокна соматической (скелетной) мышце млекопитающих (крыса) (Бакеева, 1980, Шубникова и др., 2001).
Аппарат Гольджи развит не очень сильно, также как и рибосомальная система. Необходимо только отметить, что актин и миозин синтезируются на полисомах, включающих от 15 до 75 рибосом.
В саркоплазме мышечных волокно обнаружено большое количество специфического белкового пигмента – миоглобина.Именно его количество и определяет принадлежность волокон к «красному» или «белому» типам. Также встречаются скопления гликогена(особенно его много в белых мышечных волокнах), тогда как липидных капель – наоборот мало.
Дата добавления: 2015-07-07; просмотров: 4564;