ЗРИТЕЛЬНАЯ СЕНСОРНАЯ СИСТЕМА
Зрительная сенсорная система (зрительный анализатор) является одной из самых важнейших, поскольку дает мозгу до 90% воспринимаемой информации. По сути, это основной сенсорный канал, который связывает человека с внешним миром. Благодаря зрению мы воспринимаем окружающий нас мир в объеме и красках, читаем и смотрим кино. Глазу принадлежит первостепенное значение в многообразной трудовой деятельности людей, в выполнении многочисленных весьма тонких работ. Кроме того, зрительный контроль присутствует и в определении положения тела или отдельных его частей в пространстве.
►Периферический отдел зрительной системы. Поскольку периферический отдел этого анализатора включает в себя не только рецепторы, но и целый ряд морфологических структур, обеспечивающих наиболее полное восприятие сенсорных стимулов, то необходимо хорошо представлять себе строение глаза.
Строение глаза (рис.1.1).
Диоптрический аппарат.Оптическая система глаза представляет собой сложную линзовую систему, которая формирует на сетчатке перевернутое и уменьшенное изображение внешнего мира. Диоптрический аппарат состоит из прозрачной роговицы(3),передней (4) и задней (5) камер (которые содержат так называемую «водянистую влагу»), радужной оболочки(2)(образующей зрачок), хрусталика(15),обеспечивающего аккомодацию глаза, и стекловидного тела (6), которое занимает большую часть глазного яблока. Стекловидное тело - это прозрачный гель, состоящий из внеклеточной жидкости с коллагеном и гиалуроновой кислотой в коллоидном растворе, и является оптически прозрачным.
Сетчатка.Сенсорный аппарат глаза – сетчатка(9)- развивается в процессе онтогенеза как часть промежуточного мозга. Поэтому ее называют «мозгом, вынесенным на периферию». Сетчатка покрывает внутреннюю поверхность задней части глазного яблока и состоит из нескольких слоев клеток, среди которых с точки зрения функции главными являются пигментный слой и трехнейронная цепь, состоящая из наружного – фоторецепторного (палочек и колбочек), среднего – ассоциативного, внутреннего – ганглионарного компонентов, которые пронизаны кровеносными сосудами. Выходной слой сетчатки образован из ганглиозных клеток.Их аксоны, которые вначале немиелинизированы, проходят через сетчатку к диску зрительного нерва, пересекают склеру(7)глаза в зоне решетчатой пластины и затем в совокупности образуют зрительный нерв(10). Зрительный нерв человека состоит примерно из миллиона аксонов, окруженных глиальными клетками и соединительной тканью. В этой своей части большинство аксонов миелинизировано.
Центральная ямка. В области заднего полюса глаза человека в сетчатке имеется небольшая зона, называемая центральной ямкой (13). В отличие от остальной сетчатки в этой зоне слой рецепторов не загорожен другими нейронами сетчатки (если смотреть со стороны стекловидного тела). В области центральной ямки острота зрения максимальная. В том случае, если объект фиксируется глазом, его изображение попадает в область центральной ямки.

Рис.1.1. Схема горизонтального разреза правого глаза человека
1 – конъюктива; 2 – радужная оболочка; 3 – роговица; 4 – передняя камера; 5 – задняя камера; 6 – стекловидное тело; 7 – склера; 8 – сосудистая оболочка; 9 – сетчатка; 10 – зрительный нерв; 11 – слепое пятно; 12 – желтое пятно; 13 - центральная ямка; 14 – зрительная ось; 15 – хрусталик.
Фоторецепция. Восприятие света осуществляется с участием фоторецепторов, которые относятся ко вторичночувствующим рецепторам. Все фоторецепторы (и палочки, и колбочки) состоят из двух сегментов (члеников) — наружного, светочувствительного, и внутреннего, обеспечивающего метаболизм фоторецепторов. Между собой наружный и внутренний сегменты связаны соединительной ножкой (это пучок из 16-18 тонких фибрилл).
Светочувствительные сегменты фоторецепторов обращены в сторону, противоположенную свету (наибольшее возбуждение от действия света наблюдается в тех случаях, когда направление луча совпадает с длинной осью палочки или колбочки). У палочек они представляет собой стопку тонких фоторецепторных дисков (примерно от 4000 до 1000 штук) диаметром 6 мкм каждый. Каждый диск — это двойная мембрана, состоящая из двух мономолекулярных слоев липидов, помещающихся между двумя слоями молекул белка. С этими молекулами белка связан зрительный пигмент. В палочках (их около 120 млн.) содержится только один зрительный пигмент - родопсин, в то время как в колбочках (их около 6 млн.) – йодопсин, хлоролаб, эритлаб. У палочек наружный сегмент намного длиннее и более богат зрительным пигментом, чем у колбочек. Это частично объясняет более высокую чувствительность палочки к свету: палочку может возбудить всего один квант света, а колбочку — только около 100 квантов. Фоторецепторные
диски постепенно изнашиваются (особенно интенсивно — в палочках), поэтому верхушка наружного сегмента периодически обламывается и фагоцитируется клетками пигментного слоя (в палочках это совершается в утренние и дневные часы, в колбочках — в вечерние и ночные). На замену старым дискам из внутреннего сегмента фоторецептора постоянно поступают новые диски.
Внутренний сегмент фоторецепторов содержит множество митохондрий, а также ядро, рибосомы, элементы эндоплазматической сети и комплекса Гольджи. Он является основным местом образования энергии и белков. Именно во внутреннем сегменте непрерывно совершается синтез зрительного пигмента и его упаковка в фоторецепторные диски, которые затем поступают в наружный сегмент фоторецептора для замены устаревших дисков. За 1 час образуется 3 таких диска, перемещение которых в наружный сегмент осуществляется в течение 2-3 недель.
Внутренний сегмент фоторецепторов оканчивается лентовидным образованием - пресинаптическим окончанием, в котором содержатся везикулы, наполненные глутаматом. Благодаря этому образованию сигнал с фоторецептора передается на контактирующую с ним биполярную клетку.
Принципфоторецепции. В условиях темноты фоторецепторная клетка постоянно выделяет медиатор глутамат, который гиперполяризует биполярную клетку, соединенную с ганглиозной клеткой. Поэтому в темноте биполярная клетка не возбуждает ганглиозную клетку и потенциалы действия не идут по зрительному нерву к подкорковым и корковым зрительным центрам. При попадании кванта света происходит «возбуждение» фоторецепторной клетки. Оно проявляется в том, что внутренний сегмент фоторецепторной клетки гиперполяризуется и тем самым снижает выделение медиатора (глутамата), т.е. происходит снятие тормозного влияния глутамата на биполярную клетку. Это приводит к тому, что биполярная клетка возбуждается и в свою очередь (за счет выделения возбуждающего медиатора, в роли которого выступает ацетилхолин) активирует ганглиозную клетку, благодаря чему генерируются потенциалы действия, которые по зрительному нерву достигают подкорковых и корковых зрительных центров, что в конечном итоге создает возможность для восприятия зрительного образа. Надо сказать, что в горизонтальных, амакриновых и биполярных клетках потенциалы действия не возникают. Передача сигнала происходит путем медленных изменений их мембранных потенциалов: в области синапсов освобождается медиатор в таком количестве, которое зависит от величины мембранного потенциала в терминалях, образующих синаптические окончания. Наличие всетчатке горизонтальных (они регулируют синаптическую передачу между фоторецепторами и биполярными клетками) и амакриновых клеток (регулирующих взаимодействие между биполярными клетками и ганглиозными и осуществляющими процессы латерального торможения) позволяет создать оптимальные условия для детекции светового сигнала.
►Физиология проводящих путей.Зрительная информация передается в мозг через аксоны ганглиозных клеток сетчатки, которые образуют зрительный нерв (II пара черепных нервов). Правый и левый зрительные нервы встречаются у основания мозга, где формируется их частичный перекрест, или хиазма: нервные волокна, идущие от носовых (назальных) половин обеих сетчаток, пересекаются и переходят на противоположную сторону. Такой перекрест обеспечивает каждое полушарие большого мозга информацией от обоих глаз: в затылочную долю правого полушария поступают сигналы от правых половин каждой сетчатки, а в левое полушарие — от левых половин сетчаток. Волокна, идущие от височных (темпоральных) половин каждой сетчатки, продолжают идти с той же стороны (ипсилатерально), объединяясь вместе с перекрещенным пучком аксонов из контрлатерального зрительного нерва, и образуют зрительный тракт. Зрительный тракт проецируются в ряд мозговых структур (рис. 1.2).
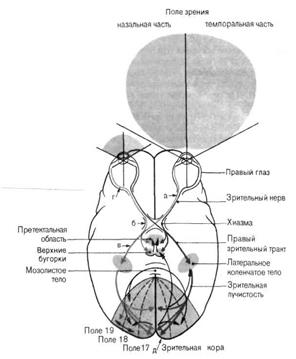
Рис. 1.2. Схема зрительных путей в головном мозгу человека.
Справа показаны эфферентные связи между зрительной корой и подкорковыми структурами. Стрелки, отмеченные буквами, указывают на возможные места, после повреждения, которых отмечаются выпадения поля зрения.
Основное число волокон приходит в таламический подкорковый зрительный центр — латеральное, или наружное, коленчатое тело. Отсюда сигналы поступают в первичную проекционную область зрительной зоны коры (17-е поле по Бродману), затем — во вторичную проекционную зону (поле 18 и 19), а затем — в ассоциативные зоны коры.
Часть волокон от ганглиозных клеток сетчатки направляется к крыше среднего мозга - к ядрам верхних бугров четверохолмия и подушку таламуса (из подушки информация передается на область 18-го и 19-го полей коры). Нейроны верхних бугров четверохолмия обеспечивают реализацию ориентировочных (сторожевых) зрительных рефлексов. Из бугров четверохолмия нервные импульсы следуют в ядра глазодвигательного нерва, иннервирующего мышцы глаза, а также ресничную мышцу и мышцу, суживающую зрачок. Благодаря этому, в ответ на попадание световых волн в глаз зрачок суживается, а глазные яблоки поворачиваются в направлении пучка света.
Часть информации от сетчатки по зрительному тракту поступает к супрахиазматическим ядрам гипоталамуса, обеспечивая тем самым реализацию циркадианных (околосуточных) биоритмов (сон – бодрствование), а также регуляцию эндокринной системы.
В передаточных станциях (верхние бугры четверохолмия, латеральные коленчатые тела) происходит вычленение зрительной информации и выявление новых качеств, недоступных «примитивной» сетчатке. С этой целью за счет явления конвергенции и дивергенции создаются более сложные рецептивные поля, а также появляются более «обученные», более «смышленые» нейроны, которые возбуждаются на особые сигналы, например, на стимул, траектория движения которого имеет волнообразный характер. В этих передаточных станциях имеет место сохранение топического расположения рецепторов на сетчатке, а также наличие колонок — вертикальных скоплений нейронов, предназначенных для расчленения информации, поступающей от данной области сетчатки, на отдельные составляющие. Например, в латеральном коленчатом теле имеются нейроны, реагирующие на свет и темноту, отвечающие за контраст, реагирующие на определенные цвета, а также на определенное направление движения светового стимула (дирекционные нейроны).
►Обработка информации в корковых зрительных центрах. Основной анализ зрительной информации совершается нейронами коры, среди которых выделяют простые, сложные, сверхсложные и гностические. Все эти нейроны объединены в вертикальные колонки (глазодоминантные, ориентационные).
Прежде всего, потенциалы действия нейронов наружного коленчатого тела поступают в затылочную часть полушарий большого мозга, где расположена первичная проекционная область зрительной зоны коры (поле 17). Афферентные волокна из наружного коленчатого тела оканчиваются в слое IV и в глубине слоя III 17-го поля, которое является центральным полем зрительной коры. К первичной проекционной зоне примыкают вторичные поля 18 и 19 (периферические поля зрительной коры). Между сетчаткой и полем 17 существует упорядоченное топографическое соответствие, благодаря чему карта сетчатки, а, следовательно, и поле зрения проецируется на кору. Центральная ямка, где острота зрения максимальна, занимает большую часть коркового представительства. Таким образом, единственную точную ретинооптическую карту содержит только поле 17.
В первичной проекционной зоне происходит анализ информации, поступающей одновременно из правого и левого глаз. Как и в других зонах коры, в этой области анализ проводится с участием колонок. Имеются глазодоминантные колонки, анализирующие информацию, идущую либо из правого глаза, либо из левого. Эти колонки соседствуют друг с другом, поэтому, вероятно, между ними происходит обмен информацией, и это позволяет видеть двумя глазами один предмет (бинокулярное зрение).
Среди нейронов колонок коры различают «простые» нейроны, задача которых состоит в выявлении контраста, наличия движущегося стимула, т. е. точно такие же задачи, как у нейронов сетчатки (но для нейронов коры рецептивные поля имеют более обобщенный характер). Простые нейроны зрительной коры в целом выполняет свои специфические функции, получая сигналы от определенного участка сетчатки и откликаясь максимальной активностью на определенную форму и ориентацию стимула.
Кроме того, колонки зрительной коры содержат «сложные» и «сверхсложные» нейроны, которые возбуждаются при наличии определенных условий, например, при движении светового сигнала слева направо или снизу вверх (дирекциональные детекторы), либо возбуждаются на определенный цвет, часть нейронов лучше всего отвечает на относительную удаленность объекта от глаз. Важно подчеркнуть, что информация о разных признаках зрительных объектов (форма, цвет, движение) обрабатывается параллельно в разных частях зрительной зоны коры больших полушарий.
Таким образом, в первичной проекционной зоне происходит более специализированная и сложная, чем в сетчатке и в латеральном коленчатом теле, переработка информации. Нейроны зрительной зоны коры имеют не круглые, а вытянутые (по горизонтали, вертикали или в одном из косых направлений) рецептивные поля небольшого размера. Благодаря этому нейроны коры способны выделять из цельного изображения отдельные фрагменты линий с той или иной ориентацией и расположением (детекторы ориентации) и избирательно на них реагировать.
Вся информация от нейронов первичного (поле 17) и вторичных (поля 18 и 19) проекционных полей коры передается в передние и задние ассоциативные зоны коры (лобные и теменно-височные области коры), где с участием более «обученных» (гностических) нейронов происходит окончательное формирование образа. В частности, в ассоциативных полях содержатся нейроны, обученные узнавать все буквы алфавита, слова, лица и т. д. Когда соответствующий нейрон «узнает» предназначенное ему для узнавания, он возбуждается, и это является физиологическим механизмом восприятия. При повреждении ассоциативных участков развиваются зрительные агнозии.
►Теории цветового зрения.Ниже будут вкратцеописаны две наиболее известные теории цветового зрения – трехкомпонентная теория цветового зрения М.В. Ломоносова, Т. Юнга, Г. Гельмогольца и теория оппонентных цветов Э. Геринга. В свое время между сторонниками каждой из этих теорий велись жаркие споры. Однако сейчас эти теории можно считать взаимно дополняющими интерпретациями цветового зрения, поскольку каждая из них дает «правильный» результат к своему уровню зрительной системы, т.е. объясняет последовательную работу двух механизмов цветовосприятия, а именно первичного (цветоприемного) механизма, связанного с функцией колбочек сетчатки, и вторичного (цветокодирующего) механизма, обусловленного обработкой зрительной информации в нейронах сетчатки, подкорковых и корковых центрах зрения.
Согласно трехкомпонентной теории, цветовое восприятие обеспечивается тремя типами колбочек с различной спектральной (цветовой) чувствительностью. Одни колбочки обладают максимумом возбуждения при их освещении красно-оранжевым (671-700 нм) светом, вторые — при освещении зеленым (546 нм) светом, а третьи — при освещении сине-фиолетовым (397-435 нм) светом. Всякий цвет оказывает действие на все три цветоприемных элемента, но в разной степени. Отметим, что до настоящего времени дискутируется вопрос о колбочках, чувствительных к зеленому цвету: по мнению ряда исследователей, они более чувствительны к желтому цвету, т.е. являются «желтоузнающими».
Разновидностью трехкомпонентной теории цветоощущения является представление о том, что все колбочки в сетчатке одинаковы и каждая из них содержит три различных светочувствительных вещества (цветоприемных элементов). Одно из них распадается при действии, главным образом, красного цвета, другое — зеленого, а третье — синего. Следовательно, в каждой колбочке имеется три приемника света и каждый из трех компонентов цветоощущения передается по своей системе сигналов (коду), отличной от других компонентов. Комбинацией излучений этих основных цветов можно получить все оттенки спектра, воспринимаемого зрением. Если одновременно и в одинаковой степени раздражаются все три типа цветоприемных элементов колбочек, то возникает ощущение белого цвета.
В целом, первый вариант трехкомпонентной теории цветоощущения подтвержден в опытах, в которых микроспектрофотометром измеряли поглощение излучений с разной длиной волны у одиночных колбочек сетчатки человека. Трехкомпонентная теория хорошо согласуется с представлением об аномалии цветовосприятия. Так, человек, имеющий все три вида колбочек, т.е. трихромат, обладает нормальным цветовосприятием. Отсутствие одного какого-либо типа колбочек (т.е. у дихроматов) приводит к нарушению цветоощущения, которое получило название частичной цветовой слепоты. Чаще это явление называют дальтонизмом — по имени известного деятеля Великой французский революции Д. Дальтона, носителя этого вида аномалии, который впервые описал ее. Дальтонизм встречается у 8% мужчин и намного реже у женщин (его возникновение связывают с отсутствием определенных генов в половой непарной у мужчин Х-хромосоме). Нарушение работы красного механизма (протанопия) встречается примерно у 1,1% мужчин, зеленого механизма (дейтеранопия) — у 1,5%, а синего механизма (тританопия) — у 0,001% мужчин. Протанопы («краснослепые») не воспринимают красный цвет, сине-голубые лучи кажутся им бесцветными; дейтеранопы («зеленослепые») не отличают зеленые цвета от темно-красных и голубых цветов, а тританопы не воспринимают лучи синего и фиолетового цвета. У 0,01% людей имеется лишь один вид колбочек (монохроматы). Крайне редко встречаются люди с полной цветовой слепотой (ахроматы). При ахромазии в результате поражения колбочкового аппарата сетчатки человек видит все предметы лишь в разных оттенках серого. Люди с частичной или полной цветовой слепотой не могут быть полноценными водителями транспорта, поскольку они не способны различать цвет огней светофоров и дорожных знаков.
Не исключено, что процессы фоторецепции в колбочках протекают по такому же типу, как в палочках. Функцию зрительного пигмента выполняют йодопсин (в наибольшей степени он поглощает свет с длиной волны 560 нм), а также хлоролаб (546 нм) и эритролаб (671 нм).
Теория оппонентных цветов была предложена в XIX в. Э. Герингом как попытка объяснить ряд феноменов цветового зрения, в том числе феномен одновременного или последовательного контраста. В частности, Э.Геринг выдвинул представление о так называемых оппонентных цветах (красный и зеленый, синий и желтый, черный и белый), связывая это с синтезом или распадом зрительных пигментов.
В настоящее время установлено, что функцию различения цветов выполняют не только колбочки сетчатки, но и нейроны сетчатки (в том числе ганглиозные и горизонтальные клетки), нейроны наружного коленчатого тела и зрительной коры. Так, показано, что в сетчатке имеются нейроны, рецептивное поле которых устроено таким образом, что в его центре находятся колбочки, воспринимающие, к примеру, красный цвет, а на периферии — колбочки, которые нечувствительны к нему. Когда луч красного цвета возбуждает колбочки, находящиеся в центре рецептивного поля, нейрон возбуждается, в результате чего возникает ощущение красного. Если свет в основном поглощается колбочками на периферии рецептивного поля, то этот нейрон не возбуждается, что создает ощущение зеленого света. Это возможно при условии существования второго варианта рецептивного поля: в центре находятся колбочки, воспринимающие зеленый цвет, тогда возбуждение этих нейронов даст ощущение зеленого, если же кванты света воспринимаются в основном колбочками периферии, то нейрон не возбуждается, что вместе с другим типом объединения рецепторов дает ощущение красного. Синий и желтый цвет возникает в рецептивных полях, где в одном случае центр представлен колбочками, воспринимающими желтый цвет, а периферия — синий, и наоборот, рецептивные поля, где в центре концентрируются колбочки, воспринимающие синий цвет, а на периферии — желтый. Ощущение черного и белого цветов возникает в результате такой организации, когда центр воспринимает все цвета (т.е. колбочки чувствительны ко всем цветам — это дает ощущение белого цвета), а периферия — не чувствительна к свету. Таким образом, любой цвет может быть представлен совокупностью нейронов, воспринимающих основные цвета. В результате многочисленного объединения нейронов на более высоком уровне (латеральное коленчатое тело, кора больших полушарий) возникает ощущение всех оттенков цвета. Оппонентность отмечена также для биполярных и некоторых горизонтальных клеток: цвет различного спектрального состава вызывает либо деполяризацию этих клеток, либо гиперполяризацию. Показано, что в наружном коленчатом теле спектрально оппонентные нейроны (цветочувствительные нейроны) представлены очень широко — на их долю приходится около 67% от общего числа нейронов. В зрительной коре также выявлены цветочувствительные нейроны. Их рецептивные поля устроены таким образом, что центр возбуждается на длинноволновый стимул и тормозится коротковолновым; напротив, периферия возбуждается коротковолновым и тормозится длинноволновым стимулом.
Таким образом, трехкомпонентная теория цветовосприятия (колбочки трех видов) достаточно хорошо согласуется с оппонентной теорией. В определенной степени эти две теории подтверждаются явлением одновременного и последовательного цветового контраста. Явление одновременного цветового контраста проявляется в том, что при длительном наблюдении серый круг приобретает зеленый цвет; если же серый круг имеет ободок зеленого цвета, то при длительном наблюдении ободок приобретает красную окраску. Явление последовательного цветового контраста (последовательных цветовых образов) проявляется в том, что если долго смотреть на окрашенный предмет (например, на красный квадрат), а затем перевести взгляд на белую бумагу, то тот же предмет виден окрашенным в дополнительный (зеленый) цвет. Причина этого явления в цветовой адаптации, т. е. снижении чувствительности к этому цвету. Поэтому из белого света как бы вычитается тот, который действовал на глаз до этого, и возникает ощущение дополнительного цвета.
Дата добавления: 2015-07-22; просмотров: 1260;