ВЕСТИБУЛЯРНАЯ СЕНСОРНАЯ СИСТЕМА
Вестибулярная система (статокинетический анализатор), взаимодействуя со зрительной и проприорецептивной сенсорными системами, играет важную роль в пространственной ориентации организма. Вестибулярному анализатору принадлежит ведущая роль в обеспечении контроля статических и динамических реакций организма, а также в определении положения тела в пространстве по отношению к гравитационному полю Земли и положения головы и шеи по отношению к туловищу. Кроме того, вестибулярный анализатор отвечает за восприятие линейных (изменение скорости движения) и угловых (перемена направления движения) ускорений. При равномерном движении или в условиях покоя вестибулорецепторы не возбуждаются.
►Строение и функции рецепторного вестибулярного аппарата.Периферический отдел вестибулярной системы - вестибулярный аппарат, который является одной из частей перепончатого лабиринта, другой его частью является орган слуха (см. Гл. 2). Аппарат состоит из преддверия, трех полукружных каналов, расположенных в трех взаимно перпендикулярных плоскостях: передний канал — в сагиттальной плоскости, задний — во фронтальной, а латеральный — в горизонтальной (рис.3.1). Такое расположение позволяет осуществлять контроль за различными поворотами головы в любой плоскости.
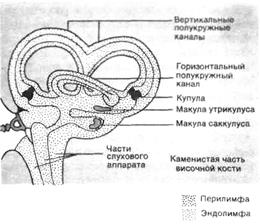
|
Рис. 3.1. Схема вестибулярного лабиринта (по Р. Шмидту и Г. Тевсу, 1985).
Лимфатические пространства вестибулярного лабиринта и улитки сообщаются между собой.
Одна из ножек каждого перепончатого канала, как и костного, расширяется и образует перепончатую ампулу (переднюю, заднюю и латеральную). Участки ампул, содержащие рецепторные волосковые клетки, называют кристами, или гребешками. В преддверии находятся два мешочка (утрикулус и саккулус), образующие отолитовый (статолитовый) аппарат. Участки этих мешочков, содержащие рецепторные волосковые клетки, называют макулами, или пятнами.В рецепторах макул и крист различают два вида ресничек – стереоцилии (по 60-80 на каждой клетке) и киноцилии (по одной на каждой клетке). И в области макул, и в полукружных каналах содержащий рецепторы сенсорный эпителий покрыт желеобразной массой. В отолитовом аппарате она называется отолитовой мембраной и содержит отложения карбоната кальция (СаСО3) в форме небольших кристаллов кальция. В полукружных каналах такая мембрана не содержит кристаллов и называется купулой. В отолитовую мембрану и купулу погружены волоски рецепторов.
Механоэлектрические преобразования на уровне вестибулярного сенсорного эпителия макул и крист. Отолитовая мембрана имеет существенно больший удельный вес (т.е. тяжелее), чем эндолимфа, заполняющая внутреннюю полость мешочков. На этом и основывается механизм действия стимулов. Так, если орган подвергается действию линейного ускорения,сила инерции, действующая на эндолимфу и отолитовую мембрану, различна, так как различна их плотность. Таким образом, весь отолитовый аппарат очень легко скользит по сенсорному эпителию подобно тому, как движутся по инерции незакрепленные предметы в резко затормозившем автомобиле. В результате реснички отклоняются и рецептор получает адекватный стимул и тем самым макулы воспринимают изменение силы тяжести линейного ускорения. Наиболее распространенной формой линейного ускорения является ускорение силы тяжести.Макулы постоянно подвергаются ее воздействию. Когда тело выпрямлено и голова находится в нормальном положении, макула утрикулуса (см. рис. 3.1) расположена почти горизонтально, так что отолитовая мембрана не прикладывает усилия сдвига к подлежащему сенсорному эпителию. Когда же голова наклоняется, макула утрикулуса оказывается расположенной под углом и тяжелая отолитовая мембрана соскальзывает на небольшое расстояние по сенсорному эпителию, в результате чего реснички изгибаются и происходит стимуляция рецептора. В зависимости от направления наклона частота импульсации в афферентных волокнах увеличивается или уменьшается. Стимуляция макулы саккулуса в принципе происходит так же, но эта структура расположена почти вертикально при нормальном положении головы (см. рис. 3.1). В невесомости отолитовая мембрана перестает давить на рецепторы, в результате чего мозг не получает информации, где верх, а где низ. Таким образом, при любом положении головы каждая из отолитовых мембран занимает определенное положение относительно рецепторов, что связано с определенной картиной возбуждения в нервных волокнах, которая в последующем обрабатывается в центральном отделе вестибулярной системы и в результате организм получает информацию о положении головы в пространстве. Это является наиболее существенной функцией отолитового аппарата.
Второй вид адекватных стимулов для ресничек вестибулярных рецепторов воспринимается в полукружных каналах (см. рис. 3.1). Последние действуют, как заполненные эндолимфой замкнутые круговые каналы, хотя их форма и не является идеально круговой. Расположенная в полукружных каналах, не имеющая минеральных включений купула обладает практически такой же плотностью,что и эндолимфа. Поэтому линейное ускорение, т.е. если голова находится в покое или во время равномерного и прямолинейного движения, не влияет на рецепторы крист. К тому же взаиморасположение каналов, купулы и ресничек при этом не изменяется. Иной эффект у углового ускорения(вращения). Так, если же голова поворачивается в сторону, то давление эндолимфы на стенки полукружных каналов меняется. Вследствие инерции эндолимфа в канале смещается и сильнее давит на сторону, противоположную движению. Воспринимающие клетки реагируют на это изменение и посылают в мозг соответствующую информацию. Следовательно, воспринимающие клетки полукружных каналов реагируют на угловое ускорение, то есть на перемену направления движения, если даже скорость его остается постоянной. Они воспринимают также вращательное ускорение и замедление.
Нервные импульсы, передаваемые волокнами, идущими от полукружных каналов, также перерабатываются в ЦНС. По картине активности нервных волокон, приходящих из трех полукружных каналов с каждой стороны, мозг получает информацию об угловом ускорении головы. Именно потому, что голова может вращаться вокруг трех пространственных осей – наклоняться вперед и назад (кивать), влево и вправо и поворачиваться вокруг длинной оси тела, - и необходимы именно три полукружных канала, расположенные в трех почти перпендикулярных друг другу плоскостях (горизонтальный расположен не совсем горизонтально: его передний край приподнят приблизительно на 300, что учитывают при клинических исследованиях).
Таким образом, адекватным раздражением для полукружных каналов являются угловые ускорения и замедления, возникающие при вращении человека или животных в горизонтальной, фронтальной или сагиттальной плоскостях, а также смена направления движения, если даже скорость его остается постоянной. Отолитовый же аппарат воспринимает как раздражения от ускорения или замедления прямолинейного движения, так и изменения в положении головы, вызывающие смещение отолитовой мембраны.
►Вестибулярные пути и обработка информации в коре больших полушарий.Первыми нейронами, воспринимающими возбуждение волосковых клеток вестибулярного аппарата, являются биполярные нейроны, составляющие основу вестибулярного, или преддверного, узла (ганглия Скарпе), который залегает на дне внутреннего слухового прохода. Их дендриты, контактирующие с волосковыми клетками, в ответ на возбуждение этих клеток-рецепторов генерируют потенциалы действия, которые передаются в ЦНС по аксонам. Аксоны биполярных клеток образуют вестибулярную, или преддверную, часть преддверно-улиткового нерва (VIII пара черепно-мозговых нервов). Физиологические исследования показали, что в вестибулярном нерве даже при полном покое наблюдается спонтанная электрическая активность. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при поворотах в другую (детекция направления движения). Реже частота разрядов повышается или, наоборот, тормозится при любом движении. У 2/3 волокон обнаруживают эффект адаптации (уменьшение частоты разрядов) во время длящегося действия углового ускорения.
Волокна вестибулярной части нерва оканчиваются главным образом в области вестибулярных ядер продолговатого мозга. С каждой стороны имеется по четыре ядра, отличающихся друг от друга как анатомически, так и функционально: верхнее (ядро Бехтерева), медиальное (ядро Швальбе), латеральное (ядро Дейтерса) и нижнее (ядро Роллера). Такая классификация основана на ответах нейронов этих ядер на натуральную стимуляцию полукружных каналов. Афферентные импульсы от вестибулярных рецепторов, приходящие в эти ядра, сами по себе не обеспечивают точной информации о положении тела в пространстве, поскольку угол поворота головы, подвижной в шейном сочленении, не зависит от положения корпуса. ЦНС должна получать информацию о положении головы относительно туловища и учитывать ее при определении положения тела в целом. Следовательно, вестибулярные ядра получают дополнительную афферентацию от шейных рецепторов (от мышц и сочленений шеи). При экспериментальном разрушении этих связей возникают такие же нарушения равновесия, как при разрушении лабиринта. Соматосенсорные афференты от других суставов также приходят в вестибулярные ядра.
Кроме того, вестибулярные ядра связаны и с различными отделами ЦНС, что служит основой для реализации вестибулярного контроля позы в результате перераспределения тонуса мышц туловища и конечностей, обеспечивающих оптимальную ориентацию тела и относительное постоянство изображения на сетчатке глаза в условиях статических и динамических воздействий. Среди таких нисходящих связей следует отметить следующие:
1) Вестибулоспинальный тракт, волокна которого направляются к α-мотонейронам спинного мозга.
2) Вестибуломозжечковый путь, который связывает вестибулярные ядра через нижние ножки мозжечка с ядрами шатра мозжечка, доставляя к ним информацию о положении тела в пространстве. Нарушение связи мозжечка и вестибулярного аппарата приводит к развитию вестибулярной атаксии, т.е. к нарушению координации движений.
3) Вестибулоокулярный путь, который связывает вестибулярные ядра с ядрами глазодвигательных нервов (III, IV и VI пары черепных нервов). Такие связи играют важную роль в регуляции движений глаз, что необходимо для сохранения стабильного изображения на сетчатке во время движения, в частности благодаря этому мы можем читать во время езды в транспорте. При возбуждении вестибулярных ядер возникают ритмические движения глаз и головы в противоположную вращению сторону, которые сменяются быстрым скачком глаз и головы обратно (нистагм глаз и головы). Это явление широко используется в клинике для оценки состояния вестибулярной системы.
4) Вестибуло-гипоталамический путь связывает вестибулярные ядра с парасимпатическими (трофотропными) нейронами гипоталамуса, находящимися, главным образом, в преоптической и передней области гипоталамуса. При возбуждении вестибулярных ядер повышается тонус парасимпатической нервной системы, в том числе тонус языкоглоточного и блуждающего нервов (IХ и Х пары черепных нервов), что вызывает при выраженном раздражении вестибулярного аппарата вестибуло-вегетативные реакции — тошноту, рвоту, головокружение, тахикардию, потоотделение, падение артериального давления и т.д. В таких случаях говорят о кинетозе (укачивание – «морская болезнь»). Это свидетельствует о том, что за счет повышения тонуса парасимпатической нервной системы вестибулярные ядра (в пределах физиологической нормы) могут принимать участие в регуляции деятельности сердечно-сосудистой системы, желудочно-кишечного тракта и других органов. У новорожденных и у людей с удаленным лабиринтом кинетозов не наблюдается.
Через некоторое время эти явления проходят и восстанавливается нормальное состояние. Повышенная склонность к болезни движения может быть уменьшена специальной тренировкой (вращение, качели) и применением ряда лекарственных средств.
Возможно, у кого-то может возникнуть вопрос – почему же таким образом реагирует наш организм при сильном возбуждении вестибулярного аппарат? Скорее всего, это связано с тем, что вестибулярная система развивалась в ходе эволюции с расчетом движения на ногах, а не в расчете на различные ускорения, возникающие в самолетах, автомобилях и т.д.
5) Лемнисковый путь— это путь, по которому информация от вестибулярных ядер поступает (после перекреста на уровне трапециевидного тела) через медиальную петлю (лемниск) к специфическим ядрам таламуса, а от них начинается прямой путь в кору больших полушарий - нижнюю часть постцентральной извилины, верхнюю и среднюю височные извилины. Кроме того, обнаружена и вторая вестибулярная зона – в моторной коре. Это обеспечивает сознательную обработку вестибулярной информации и таким образом сознательную ориентацию в пространстве.
Дата добавления: 2015-07-22; просмотров: 1813;