Обр-е желчных к-ты и их роль в переваривании жиров. Коньюгирование желч. к-т..(см.130)
 Жёлч. к-ты-эмульгаторы. В составе желчи желчные кислоты поступают в 12-перстную кишку и аллостерически активируют панкреатическую липазу.
Жёлч. к-ты-эмульгаторы. В составе желчи желчные кислоты поступают в 12-перстную кишку и аллостерически активируют панкреатическую липазу.
Жёлч. к-ты - произв.е холестерола. В организме синтез. холевая и хенодезокеихолевая к-ты. Не являются эффективными эмульгаторами. В печени эмульгирующие св-ва жёлч. к-т увеличиваются за счёт р-ции конъюг., в к-й к карбоксильной гр. жёлч. к-т присоед. таурин или глицин. Эти производные - конъюгированные жёлчные кислоты назыв.солями жёлчных кислот. Именно они служат главными эмульгаторами жиров в кишечнике. Вторичные желчные кислоты , включая дезоксихолевую кислоту и литохолевую кислоту, образуются из первичных желчных кислот в толстой кишке под действием бактерий. Литохолевая кислота всасывается значительно хуже, чем дезоксихолевая.
121. Классификация фосфолипидов и пути их биосинтеза. Значение фосфолипидов в жизнедеятельности клетки, участие их в передаче гормонального сигнала. (см.119в)
Фосфолипиды играют важную роль в струк. и ф-ции клеточ.мембр, актив. мембр. и лизосомальных ферм., проведении нерв. имп., сверт. крови, иммунологич. р-циях, процессах кл. пролиферации и регенерации тк., переносе эл. в цепи дыхат. ферм. формир. липопрот. комплексы. Они есть во всех живых клетках.
· глицерофосфолипиды— содержат остаток глицерина
фосфатидилхолин (лецитин)
фосфатидилэтаноламин (кефалин)
фосфатидилсерин
· фосфосфинголипиды — содержат остаток сфингозина
сфингомиелины
· фосфоинозитиды — содержат остаток инозитола
Центральную роль в биосинтезе фосфолипидов играют 1,2-диглицериды (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфин-гомиелинов). Цитидинтрифосфат (ЦТФ) участвует в синтезе практически всех фосфолипидов. В качестве примера рассмотрим синтез отдельных представителей фосфолипидов.
126. Особенности окисления жирных кислот с нечетным числом углеродных атомов.Жирные к-ты с нечетным числом углеводов поступают в организм с раст.пищей и морепродуктами. Их окисление происх.по обычному пути до посл.реакции, в котор.образ.пропионал-SCoA. Суть превращения пропионила сводится к его карбоксилированию, изомеризации и образов. сукцинил-SKoA, в р-циях участвуют биотин и витам.В12.

122. Химическое строение гликолипидов и их биологическая роль. Гликолипиды широко представлены в нервной ткани и мозге. Размещаются они на наружной поверхности плазматических мембран, при этом олигосахаридные цепи направлены наружу. Наиболее вероятная их функция – рецепторная. Общей частью всех гликолипидов является церамид– соединение аминоспирта сфингозина с длинноцепочечной жирной кислотой: 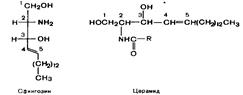

123. β-окисление ненасыщенных жирных кислот –спец.путь катаболизма жир.к-т, продуктом явл.ацетил-КоА. Окисление идет по b-углеродному атому. Служит источ.энергии для синтеза 12АТФ. Процесс идет в матриксе митохондрий в аэробных усл. Для переноса ацил-КоА ч/з мембрану митох.сущ.система – в комплексе с молекулой карнитина, ф-т карнитинацилтрансфераза I(ингибитором явл.малонил-КоА).
4реакции b-окисления наз.циклом т.к. эти 4 реакции повтор.до тех пор пока к-та не расщипится на ацетильные остатки. Регуляция: при голодании-глюкагон, в постабсорбтивном периоде-инсулин. 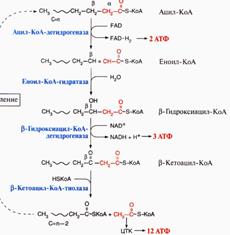
124. Окисление ненасыщенных жирных кислот, метаболические особенности этого процесса. 

125. β-окисление жирных кислот с четным и нечетным числом углеродных атомов. Энергетический выход окисления жирных кислот. жирные кислоты с нечетным числом углеродных атомов окисляются таким же образом, как и жирные кислоты с четным числом углеродных атомов, с той лишь разницей, что на последнем этапе β-окисления образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не 2 молекулы ацетил-КоА.
в сумме за один цикл образуется 5 молекул АТФ.
127. Механизм биосинтеза жирных кислот в животных клетках.митохондриальная система биосинтеза жирных кислот, включающая несколько модифицированную последовательность реакций β-окисления, осуществляет только удлинение существующих в организме среднецепочечных жирных кислот, в то время как полный биосинтез пальмитиновой кислоты из ацетил-КоА активно протекает в цитозоле, т.е. вне митохондрий, по совершенно другому пути – там идет липогенез, или de novo/ 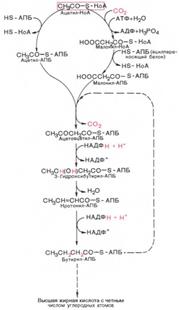
128.Биосинтез жирных кислот, последовательность реакций. Источники ацетил-КоА и НАДФН(Н)+. Локализация в клетке. См.выше.

Дата добавления: 2015-07-22; просмотров: 1126;