Регионализация и сегментация нервной трубки
Закладка telencephalon у млекопитающих контролируется парой генов Otx1 и Otx2. Эти уникальные консервативные гены млекопитающих практически не эволюционировали и эквипотенциальны гену orthodenticle (otd), контролирующему развитие переднего отдела головы дрозофилы. Мыши с двойным нокаутом Otx1-/- имели дефекты органов чувств, сенсорных систем мозга, эндокринные расстройства, развивали эпилепсию. Otx2-/- мыши погибали внутриутробно от аномалий развития переднего мозга. Поразительно, что пересадки в мозг Otx1-/- или Otx2-/- мышей гена Otd дрозофилы восстанавливали нарушенный эмбриогенез и утраченные функции переднего мозга (Acampora D., Gulisano M.,Broccoli V. et al., 2001 ). Ген дрозофилы полностью замещал функции утраченного Otx- гена млекопитающих ! Третий ген семейства Otp связан с закладкой гипоталамуса и нейро-эндокринных линий базального мозга. Otp-/- мыши погибали вскоре после рождения из-за полного отсутствия релизинг- факторов и нейропептидных гормонов гипоталамуса (Wang W., Lufkin T., 2000). Нейро-мезенхимальные взаимодействия особенно существенны на начальных фазах развития telencephalon. Нейроэпителий переднего мозга утолщается, иинвагинируя в подлежащую мезенхиму с образованием обонятельных ямок (позднее обонятельной луковицы). Ретиноевая кислота, SHH, FGF-8, BMP-4 опосредовали этот процесс. Нох-гены не принимали участие в закладке и сегментации обонятельной луковицы. Мигрирующие популяции клеток нервного гребня (НГ) составляли часть мезенхимы, формирующий обонятельную луковицу. Мутации гена Рах-6, повреждающие миграцию клеток НГ в зачаток обонятельной луковицы, приводили к аномалиям развития переднего мозга (LaMantia A.S., Bhasin N., Rhodes K. et al, 2000). Первичные культуры telencephalon 13.5 - дневных зародышей крыс использовали для изучения последовательности экспрессии второго эшелона Нох-генов. В популяции прогениторных клеток (селективно выращенных с помощью bFGF) были экспрессированы гены Otx1, Otx2, Dlx1, Dlx2, Dlx5, Emx1. Если Otx1, Otx2 маркировали популяцию некоммитированных прогениторных клеток, то Dlx /Emx маркировали постмитотические бластные линии, вступившие на путь рестрикционного созревания (Robel L., Ding M., James A.J., 1995). Пролиферацию части прогениторных клеток telencephalon контролировал FGF-8. Далее эти клетки экспрессировали BF-1 (ростовой фактор наработки клеточной массы) и Dlx2 для развития вентральных отделов telencephalon. Гомеозисный ген Vax1 контролировал развитие таламуса и визуальной системы (Hallonet M., Hollemann T., Pieler T. et al., 1999). Баланс гена-стимулятора (Vax1) и генов-репрессоров (Pax-6, Rx) задавал численность нейронов в разных отделах зрительной системы.
Гены гомеозиса (Нох-гены) разделяли территорию нервной трубки клонами некоммитированных клеток, реализующих разные soft-программы. Так, экспрессия Нох-генов маркировала границы шести прозомеров передней части трубки и семи парных ромбомеров задней части нервной трубки. Трехмерная "разбивка" территории нервной трубки устроена более сложно и зависит от множества сигналов. Сперва клетки на ранних стадиях формирования ромбомеров и прозомеров свободно мигрировали в соседние участки нервной трубки. Ограничения на миграцию возникали на более поздних стадиях развития ромбомеров. Первым барьером, намечавшим границы сегментов, был лиганд Ephrin и комплементарный ему Eph-рецептор. Клетки нейроэпителия по закону случая имели на своей поверхности либо лиганд, либо Eph-рецептор. Далее силы размежевания выстраивали вдоль границ ромбомеров слой клеток с лигандом против слоя клеток с рецептором (Wilkinson D.C., 2001 ). Данные о вкладе Нох-генов в эмбриогенез мозга получены в основном на knockout-мышах. У человека описана пока лишь одна мутация HoxD13 гена, вызывавшая полидактилию верхних и нижних конечностей.
Все 170 Нох-генов контролируют пространственные сборки зародышевых клеток первично на уровне многокомпонентных транскрипционных комплексов, куда включены не только транскриптазы, но и многочисленные кофакторы транскрипции. Поскольку кофакторы не только активируют, но и ингибируют транскрипцию, Нох-гены одновременно тормозят экспрессию «старых» генов, активируя экспрессию «новых» генов морфогенеза. Большинство мРНК Нох-генов предсинтезированы уже в созревающей яйцеклетке и предимплантационных зародышах (включая человека) (Kuliev A., Kucharenko V., Verlinsky Y. et al, 1996).
Преформированный набор мРНК сохранялся в зародыше к началу гаструляции и нейруляции. Предобразованные мРНК многих Нох-генов выявлены на стадии регионализации нервной пластинки млекопитающих (Altmann C.R., Brivanlou A.H., 2001). 39 Hox-генов кластеризовано в 4 семейства: НохА, НохВ, Нох-С и Нох-D. Эти гены экспрессированы как в трех зародышевых листках, так и мезенхиме. Мутации или делеции Hoxd3, Нох-В4, Hox-d9 , Нох-d-11 приводили к тяжелым аномалиям скелета. Парная делеция Нох-а2, Нох-а3 генов вызывала аномалии развития мышц, костно-мышечной системы лица, шеи, аномалии развития тимуса, щитовидной железы (в силу аномалий развития ромбомеров и региональных дефектов нервного гребня).
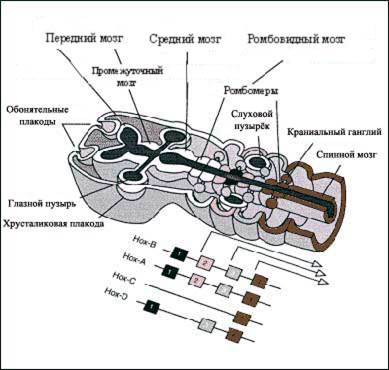
Рис 2-4. Схема действия Нох-генов в мозге зародыша млекопитающих
Зародыши мыши Lim1-/- погибали на 10-й день из-за летальных дефектов развития переднего мозга. Закладка ромбомеров заднего мозга и экспрессия Krox-20 при этом происходила нормально, что подтверждало независимость программы развития переднего и заднего мозга. Поскольку морфогенез мозга зависим от набора активных Нох-генов, это затрудняло установление вклада каждого гомеотического белка в развитие ЦНС. В ромбомерах заднего отдела мозга экспрессировались транскриптазы GATA-2, GATA-3 в субвентрикулярных слоях трубки, где локализованы НСК. Эти же Нох-гены экспрессированы в гематогенных стволовых клетках (Nardelli J., Thiesson D., Fujiwara Y. et al., 1999). Профиль активированных мРНК Нох-генов на уровне одиночных ромбомеров зародыша различен. Селекция инструкций на языке мРНК направляла формообразование в разных ромбомерах заднего мозга (Kato K., O'Dowd D.K., Fraser S.E. et al., 1997). Вторым эшелоном «регион» -сигналов трубки служили продукты гена SHH, Wnt-1, nodal и lefty, а также bFGF, BMP/GDF . Семейство генов НН состоит из трех главных генов, участвующих в регионализации ЦНС, скелетных мышц, краниофациальных структур и желудочно-кишечного тракта. В ЦНС преобладали эффекты SHH (sonic hedgehog) на общую численность и распределение региональных стволовых клеток. В других закладках регионализацию контролировали indian HH (IHH) или desert HH (DHH). Эффекты НН опосредовались двумя рецепторными трансмембранными комплексами - Patched (PTCH) и Smoothened (SMOH) ( Oldakr M., Grzela T., Lazartchuk M. et al., 2001). В постэмбриональном периоде эти гены контролировали общее количество региональных стволовых клеток в эпителии и костном мозге. У трансгенных зародышей мышей с двойной дозой SHH в нервной трубке число прогениторных клеток в клонах НСК спинного мозга увеличено в 2-3 раз ( Rowitch D.H., S-Jaques D., Lee S.M., 1999). У мышей SHH-/- возникала ранняя атрофия и недоразвитие скелетных мышц из-за резкого уменьшения численности пролиферирующих прогениторных клеток в мышечных клонах ( Kruger M., Mennerich D., Fees S., et al , 2001).
Транскриптаза Wnt-1 многократно разными путями контролировала сегментацию нервной трубки. Внутриклеточной мишенью действия Wnt-1 служит бета-катенин. Комбинируясь с разными коактиваторами, бета-катенин подключает к экспрессии новые Нох-гены. Комбинациями цитоплазматических сигналов запускается экспрессия новых Нох-генов. Модифицированные новыми белками Нох-транскрипционные комплексы направляли трехмерный рост клонов в нейромерах, ромбомерах и других обособленных территориях нервной трубки. Принято думать, что комбинации Нох-генов транслируются в направленный трехмерный рост клонов и разную численность новообразованных клеток. Рельеф доменов транскрипционных комплексов опознает "рельеф" хроматина стволовых/прогениторных клеток. Коды молекулярных соответствий переводят линейную информацию "гомеотических" генов в 3D-пролиферацию, миграцию прогениторных клеток в растущих клонах переднего, среднего и заднего мозга зародыша.
Растущий клон остается главной мишенью действия следующей батареи Нох-генов. В каждом клоне есть вентро-дорзальная и латеральная ось, вокруг которых работают градиенты продуктов Нох-генов. Появление белков Neurogenin 1 (Ngn1) и Neurogenin 2 (Ngn2) с дорзальной стороны, как и белка Mash1 c вентральной стороны нервной трубки, коррелировало с появлением первых прогениторных популяций. С этого момента доля нестин+ клеток в провизорной нервной трубке снижалась. Нокаут гена Ngn1-/- или Ngn2-/- компенсаторно увеличивал долю Mash1+ клеток в нервной ткани зародышей мышей. Баланс Ngn1+Ngn2 / Mash-1 направлял нейрогенез сенсорных/моторных нейронов спинного мозга, вентральной и дорзальной части среднего и заднего мозга. Действуя с дорзальной стороны, нейрогенины контролировали численность чувствительных нейронов, тогда как Mash-1 стимулировал созревание моторных нейронов. Сверхэкспрессия Ngn1/Ngn2 в прогениторных клетках вызывала избыточное образование нейронов не только в нервной трубке, но и в мезодерме. Нейрогенез дорзального таламуса контролировали Ngn1, Ngn2, тогда как нейрогенез вентральной части таламуса был под контролем Mash1. Далее в вентральной части нервной трубки появлялись мРНК генов семейства Nkx (Nkx 6.1, Nkx 2.2). Баланс экспрессии Mash1/Ngn направлял потоки нейро/глиогенеза в коре больших полушарий (Nieto M., Schuurmans C., Britz O, et al, 2001). У Mash1-/- эмбрионов мышей полностью блокировано образование нейронов (но не глии) в коре больших полушарий. Прогениторные клетки с экспрессией генов Mash-1 и Prox-1 теряли нестин.
Для вентро-дорзального рестрикционного созревания баланс Mash-1+ Prox-1/ Ngn1+ Ngn2 предопределял долевую численность разных линий нейронов в ЦНС (Kaibushi K., Nakamura S., Casarosa S. et al.,1999).
Отдельного рассмотрения заслуживают взаимодействия прогениторных клеток по «тандему» рецепторов Delta-Notch. Notch был открыт при изучении эмбриогенеза дрозофилы. Мутации Notch контролировали оогенез, миогенез, нейрогенез, развитие крыльев и глаза. У млекопитающих идентифицированы 4 варианта гена: Notch1-4. В нейрогенезе участвует Notch 1 и 2. Delta является внешним мембранным лигандом. Клетки с экспонированным Delta избирательно взаимодействуют с Notch рецептором других прогениторных клеток. Эта пара рецепторов стабилизирует клон за счет контактов прогениторных клеток между слоями. Эти взаимодействия, стабилизирующие клон, одновременно тормозили нейрогенез. Delta – Notch контакты прогениторных клеток в нейросферах ингибировали экспрессию нейрогенинов и Mash1, стимулируя пролиферацию. Показательно, что в растущих нейросферах эмбрионального мозга экспрессирован ген Notch-1, тогда как в растущих нейросферах постнатального мозга экспрессирован ген Notch-2. Этот маркер различал первичные и вторичные нейросферы (Higuchi M., Kiyama H., Hayakawa T. et al, 1995). Если Notch стимулировал образование новых прогениторных клеток в развивающейся ЦНС, то ген Numb отвечал за генерацию новых клонов и появление клон-инициирующих клеток. Выключение гена Numb приводило к гибели зародышей мыши на стадии 10,5-12 дня развития из-за дефектов формирования головной части нервной трубки и ускоренной генерации зрелых нейронов (Zhong W.,Jiang M.M.,Schonemann M.D. et al., 2000). Поэтому предположили, что градиент экспрессии Notch/Numb контролировал число прогениторных слоев в нейросферах и фазу перехода клона к постмитотическому созреванию (Rao M., Mattson M.P., 2001) В эпидермисе кожи человека взаимодействия Delta - Notch-1 также обеспечивали массивную экспансию прогениторных слоев клона (Lowell S.,Jones P., Le Roux et al., 2000 ). В гематогенных клонах взаимодействия Delta 1,2,3 - Notch, либо Jagged -1,2 - Notch контролировали средние размеры колоний in vitro. Взаимодействие Delta / Notch заканчивалось протеолизом цитоплазматического домена Notch. Последний транспортировался в ядро, где связывался с транскрипционным комплексом RBI-J (Schroeder T, Just U., 2000). Notch рецептор, активированный лигандом, одновременно снижал апоптоз прогениторных клеток (Han W., Ye Q, Morre M.A.,1999). Сигнализация через Delta/Notch в прогениторных клетках сопряжена с экспрессией фактора плюрипотентности Hes-1, который блокировал преждевременное включение генов Mash-1, Ngn1 Ngn2. И одновременно поддерживал уровень нестин+ и виментин+ клеток в клонах (Yuki N., Sakakibura S., Takaki M. et al., 2000).
В растущей скелетной мышце эмбрионов мышей комплексы Delta - Notch стимулировали пролиферацию некоммитированных предшественников миоцитов путем экспрессии гена Hes-1, подавляющего экспрессию гена миогенина и MyoD (Kuroda K., Tani S., Tamura K. et al., 1999). Мутации гена Notch вызывали аномалии сегментации сомитов у lunatic fringe мышей. В растущих Т-клонах, как и миелоидных клонах, Delta-Notch контролировал экспансию прогениторных популяций (Haysay A.C., Barber D.F., Douglas N. et al., 2000 ; Tan-Pertel H.T.,Walker L., Bowning D.,.2000).
Нервная плакода у зародышей млекопитающих остается наиболее древней тканью, где рекордно высока плотность Delta-Notch взаимодействий в кластерах прогениторных клеток. В интенсивно обновляющихся клонах обонятельной плакоды активированы оба фактора плюрипотентности НЕS-1 и HES-5. В клетках нейроэпителия плакоды экспрессирован только HES-1. Пока экспрессирован HES-1, невозможна экспрессия Mash-1, т.е. количество будущих нейробластов не определено в данном регионе. Пока в ткани доминировал фактор плюрипотентности HES-1, в клонах преобладала Delta-Notch зависимая пролиферация прогениторных клеток. Если HES-1 клоны определяли общую территорию, занимаемую клетками обонятельного эпителия, то ген HES-5 контролировал региональную плотность клеток (Cau E., Gradwohl G., Casasosa S. et al., 2000). В тканях мезодермы HES-7 регулировал наработку Delta-Notch прогениторных клеток, т.е. основную массу будущих мышц (Bessho Y., Miyoshi G., Sakata R. et al., 2001). Мутации (делеции) HES -генов, как правило, вели к уменьшению размеров органов эмбрионов за счет ранней, ускоренной дифференцировки прогениторных клеток и укорочения цикла HES-Delta-Notch (Kageyama R., Ohtsuka T. et al., 1999). Сам механизм размножения клонов в разных отделах головного и спинного мозга оставался неизменным, хотя пролиферация прогениторных клеток контролировалась сменными Hes- репрессорами (Ohtsuka T., Ishibashi M., Gradwohl G. et al.,1999). В цикле созревания предшественников олигодендроцитов экспрессия гена Hes -5 определяла максимальную наработку прогениторных клеток без Т3 рецептора. Выключение Hes- 5 переключало клон на наработку Mash+ прогениторных популяций, экспрессирующих Т3 рецептор. В свою очередь Т3 запускал терминальную постмитотическую дифференцировку олигодендроцитов (Kondo T., Raff M., 2000).
В ходе дифференцировки островков Лангерганса из СК дуктулярного эпителия клоны эндодермальных прогениторных клеток экспрессировали триаду генов: Delta/Notch - Math- Ngn3. На втором этапе в клетках-предшественницах островков Лангерганса включался новый набор рестрикционных сигналов: Isl-1, Brn-4, Pax-6, PDX1, Nkx6, Nkx2.2 (Schwitzgebel V.M., Scheel D.W., Conners J.R. et al., 2000).
Внутрижелудочковая имплантация эмбрионам НСК, трансфицированных Notch геном, вызывала образование радиальной глии (РГ) из клеток трансплантата. (Gaiano N., Nye J.S., Fishell G.et al., 2000). Ген Рах-6 играет незаменимую роль в закладке и развитии среднего мозга, особенно таламуса. У Рах -6-/- мышей (Sey/Sey) наблюдалась микрофтальмия и недоразвитие вентральных отделов зрительного бугра. Пересадки нормальных стволовых клеток мозга мышей в развивающиеся зародыши Sey/Sey компенсировали дефект за счет новых связй между нейронами таламуса и коры (Pratt T., Vitalis T., Warren N. et al., 2000). Пересадки нормальных донорских клеток восстанавливали экспрессию генов Nkx2.2 и Lim1/Lhx1 в вентральном таламусе. На границе среднего и заднего мозга экспрессировалось максимальное число Нох-генов: Eng-1, Eng-2, Pax-2, Pax-5, Pax-8, Pax (zf-b). Каудальнее будущего истмуса экспрессировалась другая пара генов - Wnt-1 и FGF-8. Позднее в участке трубки, кодирующей структуры заднего мозга, экспрессировались гены Еng-1 и Еng- 2, участвующие в закладке мозжечка. Мутации Eng-1-/-, Eng-2-/-, FGF-8-/- вели к остановке развития заднего мозга и гибели зародышей мышей. Нокаут Wnt-1-/- у мышей приводил к тяжелым аномалиям развития среднего мозга. Предполагается, что ген Wnt-1 контролирует транскриптазу, которая каскадным механизмом активирует Eng -1 и Eng-2 (Danelian P.S., McMahon A.P., 1996). Развитие мозжечка включало 4 этапа. Первые Нох-гены ( особенно Eng-1) размечали территорию будущего органа. На втором этапе в ромбовидной губе возникали клоны-предшественники гранулярного слоя клеток и основных ядер. Продукт гена SHH контролировал пролиферацию прогениторных клеток гранулярного слоя. Эффекты SHH частично нейтрализовались действием bFGF , либо активацией протеинкиназы А. На третьем этапе формировались основные слои мозжечка, в том числе шла миграция клеток в гранулярный слой. Секретируемый прогениторными клетками белок нетрин1 и комплементарный рецептор Unc5h3 контролируют направленную миграцию клеток как за счет сил хемопритяжения , так и хемоотталкивания (repulsion). Одна изоформа рецептора заставляет клетки мигрировать по градиенту нетрина1, тогда как другой вариант рецептора заставляет клетки избегать лиганда. Повреждение рецептора Unc5h3 или выключение гена нетрина1 приводяли к летальным аномалиям архитектоники клеток мозжечка ( Przyborski S., Knowles B., Ackerman S.,1998). Важную роль играют также два навигационных рецептора - CD10 и leu-4(CD3), которые селективным взаимодействием организуют локомоцию предшественников по волокнам Бергманновской глии (Gerloff C., Knoth R., Volk B.,1993). После рождения завершалась миграция клеток Пуркинье в гранулярный слой и заканчивалось формирование функциональных связей в нервной сети (синаптогенез).
Регулируемый сигналами апоптоз прогениторных клеток селектировал клетки для будущих сетей. До 10-го дня беременности в мозге отсутствовали погибающие клетки. На 14-й день развития до 70% прогениторных клеток элиминировались апоптозом. К 18-му дню число погибающих клеток в головном мозге снижалось до 50%. Даже в постнатальном периоде уровень репаративной обновляемости мозга остается очень высоким, поскольку 60-70% внутриутробных мозговых травм и кровоизлияний полностью компенсируется в постнатальном периоде ( Snyder E.Y., 1992). Этот же метод выявлял лишь единичные гибнущие клетки в мозге взрослой мыши ( Blaschke A.J., Staley K., Chun J., 1996). Высокий уровень апоптоза был связан с высоким уровнем сменяемости клеток клонов НСК.
Дата добавления: 2015-07-18; просмотров: 973;