Цитокинин
Впервые природный цитокинин был выделен 1964 г. Лезамом (Letham), который для этого применил многоступенчатую очистку спиртового экстракта из семян кукурузы в стадии молочной спелости [81]. Поскольку этот цитокинин был выделен из кукурузы, его назвали зеатином. Формула природного цитокинина представлена на рисунке 1.
Зеатин также был обнаружен в созревающих плодах сливы, растущих плодах яблони [82], а также в культуральной жидкости микоризных грибов [83]. В семенах люпина был обнаружен дигидрозеатин [84]. Цитокинины были также обнаружены в папоротниках, мхах и в водорослях [85].
Другое родственное зеатину вещество было выделено из культуры патогенной для растений бактерии Corynebacterium fascians. Ее действие на растения очень напоминает действие препаратов цитокинина и связано с нарушением апикального доминирования [86]. Таким образом, зеатин и родственные ему соединения с цитокининовой активностью широко распространены у представителей растительного мира.
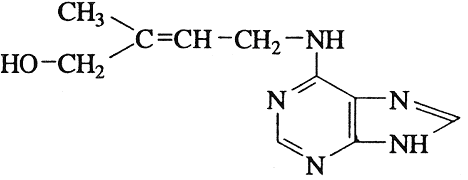
Рисунок 1 - Формула природного цитокинина – зеатина
Одним из наиболее ярких физиологических проявлений действия цитокинина является его способность задерживать старение листьев, что было впервые продемонстрирована в работе Richmond и Lang [87], в которой с помощью фитогормона удалось задержать пожелтение и распад белка в срезанных листьях дурнишника (Xanthium pensylvanicum Wallr.). В этих опытах срезанные, закончившие рост листья дурнишника помещали в раствор цитокинина. В контрольных листьях шла интенсивная убыль хлорофилла и белка, так что на 12 день опыта его потеря составляла 60% от исходного содержания. Листья на растворе фитогормона оставались зелеными и потеряли только 15% белка, что соответствовало его потере за этот срок в аналогичных листьях на растении. Таким образом, было найдено эффективное средство задержки пожелтения срезанных листьев и представлялось весьма вероятным, что цитокинины и есть те продукты корневого метаболизма, которые необходимы для поддержания нормальной жизнедеятельности листьев.
Более основательно влияние цитокинина на срезанные листья было изучено в работах [88] Mothes с сотрудниками. Ими было показано, что действие биостимулятора при его нанесении на поверхность листа остается строго локализованным в месте нанесения. Поэтому, если опрыскивать раствором цитоктинина только одну половину срезанного листа, то она сохраняется длительное время зеленой и жизнеспособной, тогда как другая половина желтеет и отмирает. Такое нанесение фитогрмона на одну половину листа было использовано в опытах на срезанных, закончивших рост листьях махорки. Эти листья сравнивали с листьями, целиком опрыснутыми водой или водным раствором цитокинина (10-20 мг/л). Все листья помещали на время опыта во влажную камеру на рассеянный свет. В контрольных листьях, получавших только воду, в течение 10 дней опыта происходило резкое снижение содержания белка и хлорофилла, которое сопровождалось оттоком азотистых соединений в черешок. Опрыскивание всего листа биостимулятором полностью предотвращало убыль белка в листовой пластинке и отток азотистых соединений в черешок. Эти результаты явились подтверждением данных Richmond и Lang [87].
Как упоминалось раньше, срезанные листья теряют способность к передвижению метаболитов из одной половины листа в другую и приобретают ее только после укоренения. Под действием цитокинина в срезанных листьях восстанавливалось передвижение метаболитов из одной половины листа в другую и, следовательно, гормон и в этом отношении как бы заменял действие корней.
Все это делало весьма вероятным предположение о том, что цитокинины являются тем самым фактором корневого метаболизма, с помощью которого корни регулируют обмен веществ листьев. Это предположение еще больше подтвердилось стимулирующим действием цитокининов на рост молодых, не закончивших рост срезанных листьев. Стимуляция цитокининами проявлялась как у этиолированных листьев в темноте [89; 90], так и у зеленых листьев на свету [91]. Причем в отдельных случаях было показано, что цитокинины могут вызвать вторичный рост уже закончивших рост листьев.
Чтобы проверить, насколько широко распространена в растительном мире способность листьев реагировать на цитокинин, Кулаева и сотрудники изучили действие кинетина на срезанные листья растений, относящихся к различным систематическим группам, подобранным в филогенетическом аспекте. Для этого срезанные листья помещали во влажную камеру на рассеянный свет при комнатной температуре и одну половину каждого листа опрыскивали водой, а другую — водным раствором кинетина (20 мг/л) [92].
Проведенное обследование показало, что кинетин замедлял пожелтение обработанной им половины листа у представителей самых различных в филогенетическом отношении семейств цветковых растений, включая наиболее древние (Ranales) и наиболее продвинутые (Glumiflorae и Campanulatae) семейства. Цитокинин влиял на листья однодольных и двудольных растений. Он действовал на листья растений, относящихся к самым различным экологическим группам, например на степное растение - ковыль и на представителя флоры влажных мест - белокрыльник. Чувствительными к кинетину оказались также и архегониаты. В частности, реакцию на кинетин проявили листья голосемянного растения Ginkgo bilobа и спорового растения - папоротника Osmunda claytoniana. Таким образом была доказана универсальность действия цитокинина на все виды растения. Интересные результаты получены в работе [93]. Была показана возможность вторичного позеленения срезанных листьев под действием цитокинина. Для опытов были взяты уже пожелтевшие на растении листья махорки. Листья срезали с растений, помещали во влажную атмосферу. Одну половину каждого опытного листа опрыскивали раствором (20 мг/л) высокоактивного цитокинина — 6-бензиламинопурина (БАП), другую - водой. Из обеих половин листа брали пробы для биохимических анализов и цитологических исследований [94; 95]. БАП вызывал вторичное позеленение обработанных им половин срезанных желтых листьев. Не обработанные БАП половины всех листьев продолжали желтеть и к концу опыта (10-11 день) большая часть их отмирала. Позеленение продолжалось в течение всего опыта. Оно шло как за счет хлорофилла а, так и хлорофилла b. Прирост хлорофилла в исходно желтых листьях шел дольше и его содержание практически достигало содержания хлорофилла в старых, но еще зеленых листьях на тех же растениях, с которых срезали опытные листья. Таким образом, обратимость пожелтения старых листьев осуществлялась тем полнее, чем менее далеко зашло пожелтение. В половине листа, получавшей только воду, происходило дальнейшее снижение содержания белка и РНК. Как отмечалось ранее, продукты распада не накапливались в желтой половине листа, а передвигались в половину, восстанавливающую свою жизнедеятельность под действием цитокинина [95; 96]. Об этом свидетельствуют данные по содержанию общего фосфора [96].
Следующая задача заключалась в том, чтобы выяснить, действительно ли корни поставляют в листья вещества цитокининового типа.
В лаборатории Mothes было показано, что срезанные листья табака обладают высокой чувствительностью к кинетину, тогда как листья на растении не дают реакцию на кинетин [97; 98]. Чтобы выяснить, какие органы растения препятствуют проявлению действия цитокинина на листья растении было проведено сравнение реакции листьев растений на фитогормон в следующих вариантах опыта: 1 - срезанный лист; 2 - лист на побеге, срезанном по корневой шейке; 3 - лист на пеньке с удаленной остальной надземной массой, но сохраненной корневой системой; 4 - лист на целом растении. Для опыта были выбраны бледно-зеленые листья третьего яруса растений махорки.
Было установлено, что фитогормон задерживает пожелтение той половины листа, на которую он нанесен. Возникающее при этом различие в пигментации двух половин листа является очень чувствительным тестом реакции листа на цитокитнин. Также было установлено, что гормон был эффективен при действии на лист находящийся на отделенном побеге, в то же время он не был эффективен при действии на лист соединенным с корневым пеньком или на лист целого растения. Таким образом, было доказано, что именно корень является продуцентом цитокинина для листьев. Аналогичные данные о важности корня как источника цитокинина были получены для растений фасоли в лаборатории Mothes [97].
Действие цитокининов проявлялось также и на различных клеточных структурах листа. Оно очень четко сказывалось, например, на внутренней структуре хлоропластов. Внутренние мембранные структуры хлоропластов являются местом локализации пигментов и ферментативных систем, обеспечивающих фотолиз воды и фотосинтетическое фосфорилирование [99]. Было показано, что под действием цитокининов дольше сохраняется структура хлоропласта [100-107].
Таким образом, установлено, что цитокинины являются одним из факторов гормональной регуляции развития и поддержания внутренней структуры хлоропластов, обеспечивающей их фотосинтетическую активность. В соответствии с приведенными данными, на срезанных листьях пшеницы, также было показано, что фитогормон задерживает разрушение в клетках хлоропластов. Точно так же он задерживал разрушение гран и заполнение хлоропластов осмиофильными глобулами в дисках из листьев брюссельской капусты [103]. Такое действие цитокининов на структуру хлоропластов должно было отражаться на их функциональном состоянии. Мембранный аппарат хлоропластов является формой пространственной организации пигментной и ферментативной систем фотосинтеза. Поэтому развитие мембран под действием цитокинина должно было приводить к активации фотосинтеза в листьях.
Дата добавления: 2015-06-27; просмотров: 2461;