БУДОВА І ВЛАСТИВОСТІ БІОЛОГІЧНИХ МЕМБРАН
Біологічні мембрани - тонкі напівпроникні оболонки, що відокремлюють клітину від зовнішнього середовища і функціональні одиниці клітини одну від одної. На рис. 4.15 схематично зображена структура клітини та її функціональних одиниць: ГлЕР - гладенький та ГрЕР - гранульований ендоплазматичні ретикулуми, ЕПС - ендоплазматична сітка, Ліз - лізосома, Ліп - ліпосома, М - мітохондрія, Мв -мікроворсинки, Мт - мікротрубочки, ПМ - плазматична мембрана, Риб - рибосоми, СГ - система Гольджі, Хр -хромосоми, Ц - центріоль, Я - ядро, я - ядречко, ЯО -ядерна оболонка.
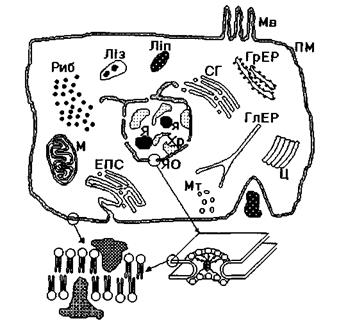
Рис. 4.15. Схематична структура клітини та її функціональних одиниць, відокремлених біологічними мембранами.
Вивчення структури і функцій біологічних мембран - сучасна область біофізики, яка має надзвичайно важливе значення для теоретичної і клінічної медицини. Порушення структури і бар'єрної функції мембран призводить до багатьох патологій, ракового переродження тканин, тканинної гіпоксії, пошкоджень, що виникають при інтоксикаціях, під впливом іонізуючої радіації тощо. Відомо, що дія багатьох лікарських препаратів спрямована на зміну властивостей та функцій саме біологічних мембран і залежить від здатності цих препаратів проникати крізь біомембрани або зв'язуватись з ними.
Головним будівельним матеріалом біомембран виступають амфіфільні (амфіпатичні) молекули фосфоліпідів - сполук з гідрофобним хвостом і гідрофільною голівкою. До складу полярної голівки, що становить приблизно одну чверть всієї довжини молекули, входить гліцерин, фосфорна кислота і полярне сполучення, характерне для кожного класу фосфоліпідів (холін, серин та інші). Гідрофобний хвіст, що становить три чверті довжини молекули фосфоліпіду, складається із залишків жирних кислот, одна з яких насичена, а інша ненасичена, з подвійними вуглецевими зв'язками (рис. 4.16).
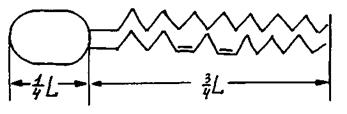
Рис. 4.16. Схематичне зображення молекули фосфоліпіду.
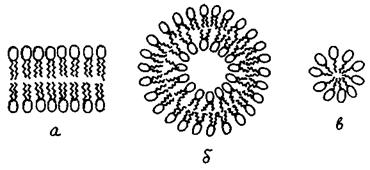
Рис. 4.17. Самозбірка фосфоліпідів у водному розчині: а - ліпідний бішар; б - везикула (ліпосома); в - міцела.
Молекули ліпідів здатні самодовільно об'єднуватись, утворюючи у воді протяжні бішарові структури (рис. 4.17а), котрі намагаються замкнутися самі на собі, ховаючи гідрофобні ділянки від води. У результаті утворюються везикули (ліпосоми) (рис. 4.17б). Процес цей називають самозбіркою. Таким чином, створення ліпідних бішарів і везикул - процес самодовільний, пов'язаний з фізико-хімічними особливостями фосфоліпідів і електростатичною дією водного оточення.
Фосфоліпідний бішар виконує функцію матриці для білків, гліколіпідів, глікопротеїдів, відіграючи водночас роль бар'єра для іонів та молекул водорозчинних речовин.
Внаслідок перекисного окислення одного з жирнокислотних ланцюгів або його відщеплення під дією ферменту фосфоліпази, може утворюватись фосфоліпід з розміром голівки, що перевищує розмір гідрофобної частини. Такі дефектні молекули утворюють не бішар, а міцелярні структури (рис. 4.17в). Опинившись у складі мембран, такі молекули утворюють пору, внутрішню поверхню якої формують полярні голівки (рис. 4.18). Крізь таку гідрофільну пору легко проходять молекули води та іони, внаслідок чого порушуються бар'єрні властивості мембрани.
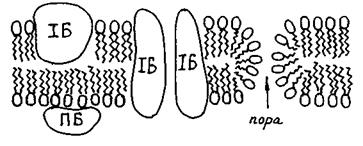
Рис. 4.18. Загальна схема будови біологічної мембрани (ПБ - периферичні білки, ІБ - інтегральні білки).
Білки мембран можуть знаходитись на поверхні ліпідного шару (периферичні білки, ПБ), утримуючись переважно електростатичними силами або вбудовуватись до ліпідного бішару, іноді пронизуючи його наскрізь (інтегральні білки, ІБ) (див. рис. 418). Інтегральні білки, опинившись поряд, можуть утворювати білковий канал (БК).
Основу сучасних уявлень про будову мембрани складає рідкокристалічна концепція, створена у 1972 році С. Сингером та Дж. Нікольсеном і вдосконалена у 1981 році С. Сингером.
Рідкі кристали - це особливий стан деяких речовин, переважно органічних, якому притаманна плинність (як рідині), але молекули при цьому зберігають впорядкованість у розташуванні, що спонукає до анізотропії ряду фізичних властивостей (як у кристалів). Згідно з цим уявленням, бішар - це рідка структура, в якій молекули ліпідів здатні здійснювати сегментальну рухливість, обертальні рухи і латеральну дифузію, що являє собою послідовний обмін місцями у межах одного шару (рис. 4.19).
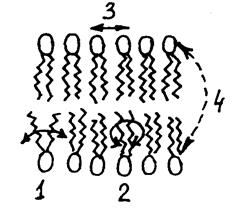
Рис.4.19 Різні види рухомості ліпідів в бішарі: 1 - сегмен- тальна рухливість; 2 - обертальна рухливість; 3 - латеральна рухливість; 4 – перехід 
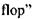
Оцінимо частоти перескоків 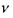 і характерні відстані таких перескоків при латеральній дифузії. Час перескоку оцінюється за формулою
і характерні відстані таких перескоків при латеральній дифузії. Час перескоку оцінюється за формулою 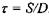 , де
, де 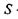 - площа, яку займає одна фосфоліпідна молекула
- площа, яку займає одна фосфоліпідна молекула 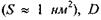 - коефіцієнт латеральної дифузії
- коефіцієнт латеральної дифузії 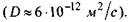 Згідно з цією формулою,
Згідно з цією формулою, 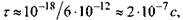 а частота перескоків
а частота перескоків 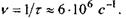 Середню відстань при перескоці оцінимо за формулою
Середню відстань при перескоці оцінимо за формулою 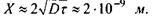 Тоді за рахунок латеральної дифузії молекула ліпіду пройде за час
Тоді за рахунок латеральної дифузії молекула ліпіду пройде за час 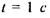 відстань
відстань  тобто досить велику відстань. Такий розмір має бактерія типу Е.соlі.
тобто досить велику відстань. Такий розмір має бактерія типу Е.соlі.
Із значно меншою швидкістю молекули здатні здійснювати перехід на інший бік бішару (перехід  , див. рис. 4.19). Цей перехід відбувається зі швидкістю одна молекула за декілька годин. Білки в ліпідному бішарі також досить рухливі. Період обертального руху білка в бішарі становить
, див. рис. 4.19). Цей перехід відбувається зі швидкістю одна молекула за декілька годин. Білки в ліпідному бішарі також досить рухливі. Період обертального руху білка в бішарі становить  Щодо латеральної рухливості білка, то вона визначається не тільки його властивостями, але й мікров'язкістю ліпідного оточення, тобто фазовим станом ліпідного бішару.
Щодо латеральної рухливості білка, то вона визначається не тільки його властивостями, але й мікров'язкістю ліпідного оточення, тобто фазовим станом ліпідного бішару.
Фізичні методи досліджень свідчать про те, що ліпідний бішар може перебувати у двох фазових станах: 1) у стані твердого двомірного кристалу; 2) у рідкокристалічному стані. При фазовому переході змінюється рухливість полярних груп, а також обертальна рухливість С-С зв'язків вуглеводневих ланцюгів (рис. 4.20).
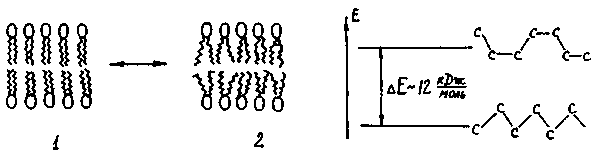
Рис. 4.20. Фазовий перехід у Рис. 4.21. Конформаційні пере-мембрані: 1 - "твердий" стан, 2 - творення ліпідів при фазовому "рідкий" стан. переході.
Якщо мембрана знаходиться у твердій фазі, то в ній існують ліпіди лише у стані трансконформерів (рис. 4.21). У рідкій фазі з'являються транс-гош-переходи. Після плавлення бішар становить динамічну суміш транс - та гош-конформерів. Гош-конформери, розташовані поряд, утворюють порожнини в бішарі чи так звані "кінки". Синхронні транс-гош переходи можуть бути представлені як рух "кінків" вздовж вуглеводневих ланцюгів. Разом з цим рухом крізь мембрану можуть проникати молекули гідрофільних речовин, уникаючи пори та канали (рис. 4.22).
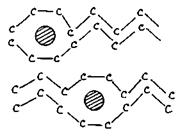
Рис.4.22. Переніс молекули внаслідок дифузії „кінка”.
Не слід думати, що вся поверхня мембрани клітин має одні і ті самі властивості. І за ліпідним складом, і за включенням білків мембрани є гетерогенними. Це також означає, що у мембранах спостерігаються дільниці з різною рухливістю (як рідкі, так і більш структуровані фази). Розділ фаз може відбуватись під впливом багатьох факторів (зокрема, внаслідок зміни 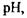 появи іонів
появи іонів 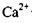 тощо).
тощо).
Кооперативна реакція типу фазового переходу - важлива властивість ліпідів, бо саме на ній базуються механізми відбирання іонів із середовища, посилення зовнішнього сигналу, мембранної пам'яті клітини. Доцільно навести деякі фізичні характеристики біологічних мембран: товщина 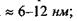 поверхнева ємність
поверхнева ємність 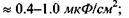 напруга в спокої
напруга в спокої 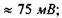 напруга пробою
напруга пробою 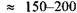
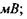 напруженість електричного поля
напруженість електричного поля 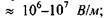 поверневий натяг
поверневий натяг 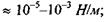 в'язкість
в'язкість 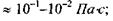
оптичний показник заломлення 
Дата добавления: 2015-06-22; просмотров: 2084;