Анаэробные термофильные бактерии
Значительную часть спороносных термофильных бактерий составляют анаэробные виды. Известны облигатно-термофильные масляно-кислые, целлюлозные, десульфурирующие и метанобразующие бактерии.
Термофильные целлюлозные бактерии. Этих бактерий обнаруживают обычно в компостах, разлагающихся растительных отбросах, илах и т. д. В таких субстратах эти бактерии наряду с термофильными целлюлозоразлагающими грибами и актиномицетами находят благоприятные условия для своего развития. Когда температура поднимается до 60—65 °С, физиологическая активность грибов и актиномицетов резко снижается и разрушение клетчатки полностью осуществляется бактериями.
Развитие целлюлозных бактерий можно заметить сначала по газообразованию, затем по разрушению клетчатки (фильтровальной бумаги) и появлению желто-оранжевого пигмента.
Чистые культуры этих бактерий получить довольно трудно, и это удавалось немногим исследователям. Типичным мезофильным представителем является бактеррш Bacillus omeli-anskii, названная в честь В. Л. Омелянского, впервые описавшего этот микроорганизм. В качестве типичного термофильного представителя можно назвать Clostridium thermocellulaseum. Описания мезофильных и термофильных видов указанных бактерий тождественны, поэтому А. А. Имшенецкий считает, что они представляют один вид. При этом термофилы могут рассматриваться как варианты мезофилов.
Морфологически целлюлозные бактерии представляют собой тонкие, прямые или слегка изогнутые палочки, часто с округлыми спорами на концах клеток. Палочки подвижны, жгутики расположены по всей поверхности клетки.
Продуктами обмена веществ анаэробных целлюлозных бактерий, выделяющимися в окружающую среду, являются водород, углекислый газ, этиловый спирт, глицерин, муравьиная, уксусная, молочная, янтарная, яблочная и фумаровая кислоты. Присутствуют также следы ацетальдегида, иногда — масляная кислота. Эти бактерии разлагают целлюлозу до низкомолекулярных углеводов, главным образом до целлобиозы и глюкозы. Гидролиз целлюлозы сопровождается появлением в среде фермента целлюлазы.
Термофильные метановые бактерии могут сопутствовать анаэробным целлюлозным бактериям или культивироваться совместно с ними. В начале этого века Ц. К о-олхаасом впервые были описаны термофильные метановые бактерии с температурными границами развития 45—69 °С. Наиболее типичными видами являются Methanobacterium soehngenii и Methanobacillus omelianskii. Эти бактерии представляют собой тонкие, прямые или слегка изогнутые неспороносные палочки.
Термофильные десульфурирующие бактерии нередко сопутствуют термофильным целлюлозным бактериям. При высоких температурах восстановление сульфатов осуществляется бактерией Desulfotomaculum nigrificans, которая представляет собой палочки с округлыми концами, иногда чечевицевидные и вздутые, подвижные, пери-трихи. Споры овальные, субтерминальные или терминальные.
Термофильные лучистые грибки (актиномицеты)
К термофильным лучистым грибкам относятся актиномицеты различных систематических групп, обладающие способностью развиваться при высоких температурах (50—60 °С), независимо от температурного минимума их роста.
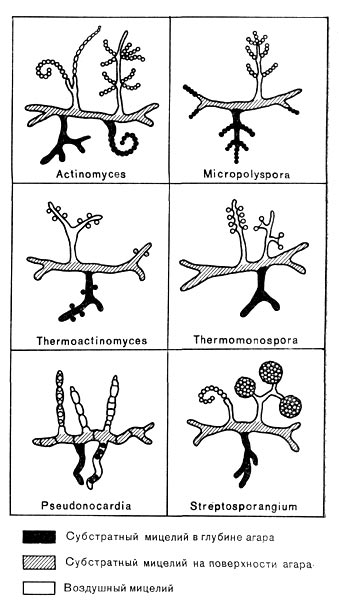
Рис. 190. Схематическое изображение вегетативного и спороносного мицелия различных родов термофильных актиномпцетов.
Среди них встречаются актиномицеты, способные расти при 60—70 °С. Лучистые грибки, развивающиеся при обычных температурах (25 — 30 °С), не растут при температуре 50 °С и выше.
Количественный учет термофильных актино-мицетов в почвах и компостах был проведен В. Ваксманом с сотрудниками в 1939 г. Термофильные актиномицеты были обнаружены во всех почвах и во все сезоны года. Особенно много их в почвах, удобренных навозом (в среднем 200 000 на 1 г в весенних и летних пробах). Зимой термофильные актиномицеты составляли 10—15% от всей термофильной микрофлоры; весной и летом 70—90%. Количество термофильных лучистых грибков не зависит от географической закономерности, а определяется экологическими факторами, в частности типом почвы и степенью ее окультуреннооти.
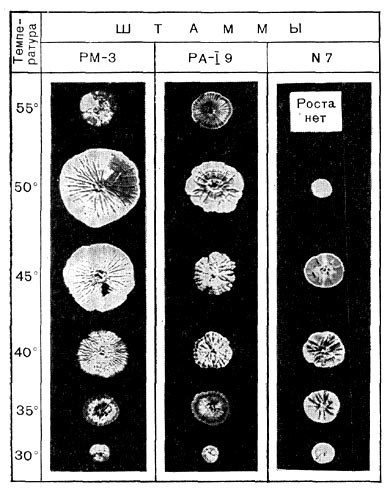
Рис. 191. Гигантские колонии различных актиномицетов при разных температурах. Уменьшено в 2,5 раза.
Н. Д. Красильников (1970) указывает, что в пределах одного семейства и рода (например, Micromonospora) могут быть как термофильные, так и мезофильные культуры; несмотря на довольно стойкий признак термофильности, видимо, нецелесообразно ориентироваться на него при характеристике родов или более крупных таксономических единиц.
Актиномицеты, способные развиваться при 40—60 СС, встречаются среди представителей различных родов, но чаще всего среди культур рода Micromonospora. Ранее признак термофильности отражали в некоторых родовых названиях актиномицетов, растущих при повышенных температурах (рис. 190). При идентификации термофильных актиномицетов используют, как обычно, культурал.ьные признаки.
Большинство известных термофильных лучистых грибков быстро гидролизуют крахмал, свертывают и пептонизируют молоко, разжижают желатин и т. д., что свидетельствует о высокой ферментативной активности и может быть использовано в практике. Однако эти культуральные свойства лабильны и поэтому, с точки зрения многих исследователей, не могут быть основными критериями при определении вида.
Другие культуральные свойства, такие, как восстановление нитратов, образование сероводорода, в большей степени отражают физиологические особенности микроорганизма, более стабильны и, следовательно, играют существенную роль при идентификации вообще и термофильных актиномицетов в частности.
Для выделения термофильных актиномицетов разными авторами использовались разные методики. Выделение этих микроорганизмов хорошо удается на крахмальном агаре, крах-мально-аммиачно-сульфатпом агаре, а также мясо-пептонном агаре (МПА). Лучшим для этой цели оказался крахмально-аммиачно-сульфатный агар, на котором наблюдается слабый рост более требовательных к среде термофильных бактерий и, наоборот, хороший рост термофильных актиномицетов. Наиболее подходящая температура для выделения 55—60 °С.
Культивируют термофильные актиномицеты на самых различных средах.
Для получения хорошего роста и споруляции этих микроорганизмов обычно используют крахмал и неочищенную мальтозу. К. Е. Эриксон отмечает, что Micromonospora vulgaris хорошо растет при использовании следующих источников азота: пептона, трип-тона, гидролизата, казеина. Многие термофильные актиномицеты нуждаются в дополнительных компонентах среды, представляющих собой смеси аминокислот, витаминов, пуринов и пиримидинов.
Термофильные актиномицеты обладают большой скоростью роста. Их жизненный цикл проходит гораздо быстрее, чем у мезофильных штаммов. Термофильные актиномицеты образуют разветвленный мицелий из гиф, диаметр которых от 0,2 до 1 мкм. На твердых средах они растут в виде плоских колоний, достигающих 3—4 см в диаметре, а иногда и до 6—8 см (рис. 191), часто покрытых налетом, состоящим из воздушных гиф со спорами. Термофильные актиномицеты образуют воздушный и субстратный мицелии. Гифы воздушного мицелия без спор термофильных лучистых грибков, как правило, белоснежно-белого цвета. Воздушный мицелий со спорами или сохраняет белый цвет, или приобретает темно-серый оттенок. Серо-зеленые, голубые и желтые штаммы встречаются реже.
У некоторых термофильных актиномицетов в процессе развития изменяется цвет колоний на агаре от белоснежно-белого до желтого, грязно-зеленоватого, коричневого, красноватого и даже черного. Многие представители термофильных лучистых грибков образуют растворимый пигмент, который проникает в среду и окрашивает ее в яркие цвета.
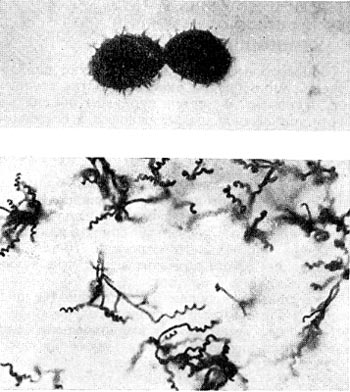
Рис. 192. Термофильный актиномпцет Actinomyces diastaticus, штамм 7. Вверху — споры с шиповидной поверхностью (увел. X 10 000); внизу - форма спороносцев (увел. X 900).
Разные исследователи описывают наличие у термофильных актиномицетов прямых или спирально закрученных спороносцев (рис. 192). Имеются также формы с одиночными, парными спорами и спорами в виде цепочек, образованных на коротких веточках вегетативного воздушного мицелия. Споры всех термофильных актиномицетов круглые или эллипсоидальные (рис. 192) диаметром от 0,6 до 1,4 мкм. В них обнаружена дипиколиновая кислота, а также кальций и в меньшей степени магний, которые, как известно, обусловливают высокую термоустойчивость спор бактерий.
Изучение тонкого строения спор ряда актиномицетов позволило исследователям сделать вывод, что некоторые термофильные актиномицеты образуют споры, близкие по строению к спорам бактерий родов Bacillus и Clostridium.
Обмен веществ у термофильных микроорганизмов происходит более интенсивно, чем у мезофильных. Об этом свидетельствуют экспериментальные данные о более высокой ферментативной активности термофилов и о повышенном количественном содержании в клетках термофильных микроорганизмов некоторых ферментов.
Так, при изучении цитохромов в клетках мезофильных и термофильных представителей различных родов актиномицетов было обнаружено, что у некоторых термофильных актиномицетов (Thermoactinomyces sp., Actinobi-fida dichotomica, Mycropolyspora sp.) оказалось больше цитохромов типов с и а; очевидно, некоторые участки цепи переноса электронов (в частности, цитохромная система) термофильных штаммов могут значительно отличаться от таковых у мезофильных форм.
Нередки случаи, когда термофильные актиномицеты способны образовывать в больших количествах экзоферменты, действующие на различные субстраты, что может быть использовано в практике. Так, выделен термофильный актиномицет Act. diastaticus штамм 7 (рис. 192), образующий целлюлозолитические и гемицеллюлозолитические ферменты. Однако образование подобных экзоферментов не является свойством, специфическим для термофилов.
С целью выяснения биохимических особенностей термофильных актиномицетов многие исследователи изучали состав нуклеотидов ДНК как термофильных, так и мезофильных видов. Результаты исследований дают возможность полагать, что термофильные актиномицеты содержат сравнительно меньше ГЦ (гуанин + цитозин) в составе ДНК, чем мезофиль-ные штаммы. Этот вопрос требует дальнейших исследований. В биомассе актиномицетов, выросших при высокой температуре (55—57 °С), содержится в 2 раза больше свободных нуклеотидов; количество нуклеиновых кислот, наоборот, падает. Свободные нуклеотиды, видимо, играют более значительную роль в интенсификации обмена веществ у термофилов по сравнению с мезофильными микроорганизмами.
Есть данные о том, что термофильные штаммы рода Actinomyces не чувствительны к фагам, активным против мезофильных штаммов того же рода, хотя существенных биохимических различий в составе клеточных стенок у мезофильных и термофильных актиномицетов не обнаружено.
Дата добавления: 2015-06-12; просмотров: 2971;