Строение лучистых грибков
Все высшие формы лучистых грибков имеют хорошо развитый мицелий. Толщина гиф мицелия варьирует в широких пределах в зависимости от видовой принадлежности культуры, ее возраста, условий развития. Диаметр тонких гиф 0,1 —1,0 мкм, наиболее толстых гиф — до 1,5 мкм. У актиномицетов, выращенных на плотных агаризованных средах (поверхностные культуры), различают три типа мицелия: субстратный, воздушный и надсубстратный, или колонии. Субстратный мицелий развивается в глубине среды и представляет собой как бы корневую питательную систему. Нити его всасывают питательные вещества среды и доставляют колонии, а оттуда в воздушный мицелий. Мицелий надсубстратный развивается на поверхности агаризованной среды, разрастается плотным сплетением в колонии большей или меньшей величины. На рисунке 83 показаны некоторые типы колоний актиномицетов. Колонии лучистых грибков плотные, кожистые, крепко срастающиеся со средой. Величина их зависит от вида актиномицетов и условий их роста, они могут быть мелкими — 0,5—2 мм и крупными — до I см и более. Внешний вид колоний также различен: колонии бывают с гладкой, бугристой, складчатой и зернистой поверхностью. Воздушный мицелий актиномицетов формируется на поверхности колонии. Нити его отходят от мицелия колонии, разрастаются в густую пушистую, бархатистую или мучнистую массу. Нередко воздушный мицелий развивается слабо в виде небольшого налета, а некоторые организмы и вовсе лишены его.
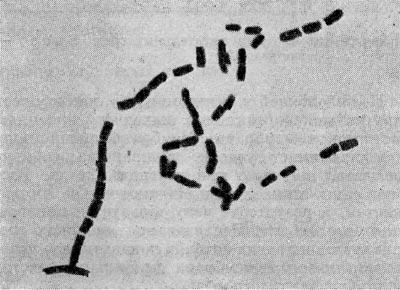
Рис. 86. Сегментацпонный способ образования спор.
Исследованиями последних лет было показано, что поверхность клеток воздушного мицелия отличается от поверхности субстратного мицелия. На поверхности клеток воздушного мицелия обнаружен рыхло связанный слой, построенный из очень тонких нитевидных частиц и различных фибриллярных телец. Субстратный мицелий называют первичным, а воздушный — вторичным. Есть сообщения и о том, что способность образования воздушного мицелия у актиномицетов может быть стимулирована или подавлена при добавлении в питательную среду определенных веществ.
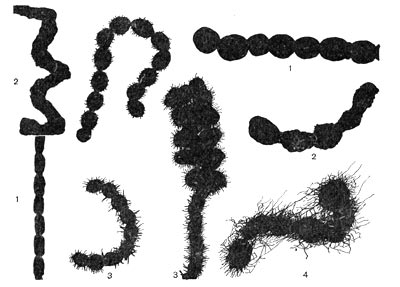
Рис. 87. Поверхность оболочки спор у актиномицетов: 1 — гладкая; 2 — бугристая; 3 — шиповидная; 4 — волосистая.
Исследователи наблюдали случаи, когда естественная потеря способности формировать воздушный мицелий у некоторых актиномицетов восстанавливается при добавке в питательную среду витаминов (биотин), аминокислот (метио-нин) и других веществ.
Клетки актиномицетов состоят из нескольких основных частей. На рисунке 84 показан общий план строения клетки актиномицетов. Клетка ограничена оболочкой — стенкой, толщина которой от 0,01 до 0,03 мкм. На поверхности оболочек мицелия расположена слизистая капсула. Обычно она очень тонкая и не обнаруживается при микроскопическом анализе, но у отдельных представителей лучистых грибков наблюдается обильное, видимое простым глазом скопление слизи, которое является специфическим признаком определенных групп организмов.
Оболочка актиномицетов в целом представляет собой относительно неплотную систему. Через нее в клетку могут проникать многие химические соединения, например белок, антибиотики, аминокислоты и многие другие вещества.
Оболочка принимает участие в общем обмене веществ и образовании различных продуктов жизнедеятельности клетки — метаболитов. Оболочка построена так же, как и у грамположи-тельных бактерий, и состоит из белковых, ли-пидных, мукополисахаридных' компонентов. У отдельных групп актиномицетов в оболочках содержатся фосфорные соединения, тейхоевые кислоты, количество которых у разных представителей актиномицетов не одинаковое.
Многие исследователи пытаются подразделить организмы на группы в зависимости от химического состава оболочек. Так, американские микробиологи лучистые грибки делят на 6 групп в зависимости от содержания в оболочках разных стереоизомеров диаминопимелино-вой кислоты (ДАП), аминокислот и Сахаров. Другие ученые считают, что пока нет достаточных экспериментальных сведений, которые позволили бы делить организмы на соответствующие группы по химическому составу их оболочек. Состав оболочек, по-видимому, не может служить единственным критерием для разделения актиномицетов, так как у одной и той же группы культур в оболочках находят различные по составу компоненты.
Под оболочкой у актиномицетов, как и у всех других микроорганизмов, находится цитоплаз-матическая мембрана. Мембрана актиномицетов принимает участие в процессах обмена веществ, а также при делении клеток. Рост мембраны приводит к впячиванию ее внутрь клеток, в результате чего образуется поперечная перегородка. Исследования японских специалистов и других авторов показали, что цито-плазматическая мембрана принимает участие и в образовании спор.
Цитоплазма клетки состоит из оптически плотных скоплений различных размеров, рисунка и сложности. В цитоплазме имеется также нуклеоид, заполненный сетью переплетенных нитей ядерного вещества. Ядерное вещество не имеет мембраны и не отделено от цитоплазмы. В старых культурах цитоплазма мицелия приобретает грубозернистую структуру, распадается на отдельные комочки. Нити одной и той же культуры не равноценны по состоянию цитоплазмы и составу оболочки. В цитоплазме актиномицетов обнаружены растворимые и нерастворимые полифосфаты, полисахариды, а у отдельных культур — жировые вещества. При ультрамикроскопическом анализе клеток лучистых грибков в цитоплазме выявляются особые структуры мезосомы с тонким мембранным строением. Наблюдается связь этих структур с цитоплазматической мембраной. Эти мембранные тельца расположены или беспорядочно, или в виде концентрических колец, рыхлые либо плотно сжатые, в одних клетках их больше, в других — меньше. Во внутрицитоплазматических мембранах концентрируются окислительные ферменты.
При спорообразовании в клетках наблюдаются следующие изменения: хроматиновое вещество разделяется на комочки различной формы и величины, располагающиеся вдоль спороносной гифы, вокруг них концентрируется цитоплазма. Затем вся цитоплазма расчленяется на участки — проспоры, которые покрываются собственной оболочкой и затем превращаются в споры.
В нитях актиномицетов, кроме хроматиновых зерен, имеются гранулы волютина, который образуется и накапливается в зависимости от состава среды, условий роста и возраста культур. Появление волютиновых гранул в цитоплазме клеток актиномицетов сопровождается резким снижением антибиотической продуктивности мицелия.
Волютиновые гранулы содержат РНК и кислотонерастворимые полифосфаты. Причем состав волютина гиф разного возраста неодинаков: на ранних стадиях развития культур в волютиновых гранулах содержится большее количество РНК, на более поздних стадиях основными компонентами волютина являются полифосфаты. Волютин — не обязательный структурный элемент клеток актиномицетов. Волютиновые гранулы появляются обычно на более поздних стадиях развития культуры в условиях нарушенного баланса между отдельными компонентами питательной среды. При культивировании актиномицетов на некоторых средах (особенно при глубинном выращивании) в цитоплазме гиф появляются вакуоли в виде маленьких или крупных пузырьков. Содержимое вакуолей в молодых гифах однородное. В зрелых нитях мицелия встречаются крупные зернистые вакуоли. Предполагается, что эти вакуоли образуются из запасных веществ. Клетки мицелия представителей лучистых грибков хорошо окрашиваются основными анилиновыми красками: метиленовой синью, метиловым фиолетовым, карболовым фуксином и др.
Характерной особенностью всех лучистых грибков является их способность окрашиваться по Граму положительно. Клетки лучистых грибков устойчивы к щелочным условиям среды и действию протеолитических ферментов (трипсина, пепсина). Грамположительные формы микробов более чувствительны к действию антибиотиков, чем грамотрицательные, и менее чувствительны к реактивам: анилину, фенолу, этиловому спирту, бензолу, хлороформу и др.
У разных представителей лучистых грибков способы размножения различны. Актиномицеты имеют специальные спороносные органы - спороносцы, или спорангии. Спороносцы у актиномицетов различаются по своему строению и расположению: прямые, длинные или короткие, волнистые; спирально закрученные; число завитков от 1 до 10 и более. Завиток может быть растянутым или плотно сжатым. Расположение спороносных веток моноподиальное (последовательное), мутовчатое и пучкообразное, супротивное. В мутовках спороносные ветки могут быть прямыми и спиральными. На рисунке 83 показаны основные типы строения спороносцев актиномицетов.
Для актиномицетов характерно образование спор — конидий. Споры формируются на воздушном мицелии, в отдельных случаях — на субстратном мицелии. Отмечено два типа образования спор — фрагментационное и сегментационное.
Фрагментационный процесс характеризуется тем, что вокруг ядерных элементов, равномерно распределенных по гифе, концентрируется цитоплазма, которая покрывается собственной оболочкой и превращается в зрелую спору. Оболочка спороносных нитей некоторое время сохраняется, затем распадается, споры высыпаются и в соответствующих условиях снова прорастают, образуя гифы новых колоний актиномицетов (рис. 85). При сегментационном образовании спор наблюдается простое деление спороносных клеток. На всем протяжении ветки образуются поперечные перегородки,и спороносец расчленяется на ряд равномерных клеток — спор. Перед сегментацией в спороносящей ветке происходит деление ядерного вещества, образуется столько зерен хроматина, сколько будет спор. Вокруг хроматиновых зерен также концентрируется плазма клетки. Когда споры созревают, спороносец распадается на отдельные споры (рис. 86).
Споры актиномицетов различаются по форме. Они бывают шаровидными, цилиндрическими, палочковидными, грушевидными. У одних видов актиномицетов поверхность оболочки спор имеет шиловидную, бугристую, волосовидную структуры, у других — поверхность оболочки гладкая (рис. 87). Этот признак используется для определения принадлежности актиномицетов к той или иной группе.
По сравнению с вегетативными клетками споры обладают повышенной устойчивостью к действию неблагоприятных факторов. Так, культуры актиномицетов, высушенные до воздушно-сухого состояния, могут сохранять жизнеспособность в течение нескольких лет. Споры устойчивы и к механическому воздействию: при растирании культур в смеси с частицами песка споры не теряют жизнеспособности, тогда как соответствующие им по размеру фрагменты вегетативного мицелия погибают. Имеются сведения, что споры актиномицетов могут свободно переноситься по воздуху. Прорастание спор у лучистых грибков однотипное. На свежей питательной среде они сначала несколько разбухают, затем на поверхности появляются бугорки — почки, которые, удлиняясь, прорастают в длинные нити. Нити ветвятся, формируя мицелий — колонии лучистых грибков.
Актиномицеты могут размножаться также обрывками мицелия разной длины, от едва заметного комочка до палочковидных элементов. Культуры актиномицетов пересеваются с помощью комочков субстратного или воздушного мицелия или самой колонии. Во всех случаях вырастают нормальные клетки дочернего организма, свойственные данному виду.
Группы лучистых грибков отличаются по способу образования органов плодоношения, что служит признаком для деления их на семейства, роды.
Дата добавления: 2015-06-12; просмотров: 1034;