Физиологические и биохимические особенности спорообразующих анаэробных бактерий
Главные отличия анаэробов касаются энергетического метаболизма, т. е. реакций, снабжающих организм энергией, и ферментов, связанных с этими реакциями. Сущность анаэробиоза была кратко и в то же время точно сформулирована Луи Пастером как «жизнь в отсутствие воздуха», т. е. жизнь без дыхания, существующая за счет анаэробного превращения веществ.
Анаэробиоз свойствен и факультативно анаэробным микроорганизмам. В отличие от последних облигатные анаэробы не могут развиваться в присутствии кислорода, более того, кислород в молекулярной форме ядовит для анаэробов.
Как известно, микроорганизмы черпают энергию, необходимую для поддержания их жизнедеятельности, за счет различного рода процессов окисления органических (а иногда и неорганических) веществ. При этом окисление происходит путем отщепления от субстратов водорода (или электронов). Водород переносится по цепи ферментов и в конечном итоге соединяется с кислородом, образуя воду. Анаэробный же способ извлечения энергии характеризуется тем, что свободный кислород в нем не принимает участия, а органические субстраты окисляются только за счет отщепления водорода. Освобождающийся водород либо присоединяется к продуктам распада того же самого органического вещества, либо выделяется в газообразном состоянии.
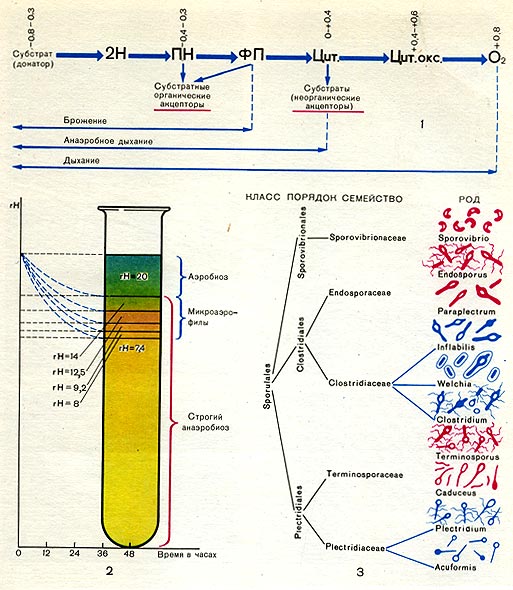
Таблица 36. Спорообразующие анаэробные бактерии.
На схеме 1 (табл. 36) показан транспорт электронов при дыхании и различных типах анаэробного способа получения энергии. Водород и электроны отщепляются от субстратов с помощью пиридиннуклеотидных ферментов (ПН). Далее они у аэробов проходят по цепи ферментов с возрастающими потенциалами — флавопротеидные (ФП)—цитохромные ферменты (Цит.) - и с помощью цитохромоксидазы (Цит. оке.) переносятся на кислород. Поток электронов направлен от системы с более низким (более отрицательным потенциалом) к системе с более высоким (более положительным) потенциалом, от — 0,8 — 0,4 в (потенциал субстрата) до + 0,8 в (потенциал кислорода).
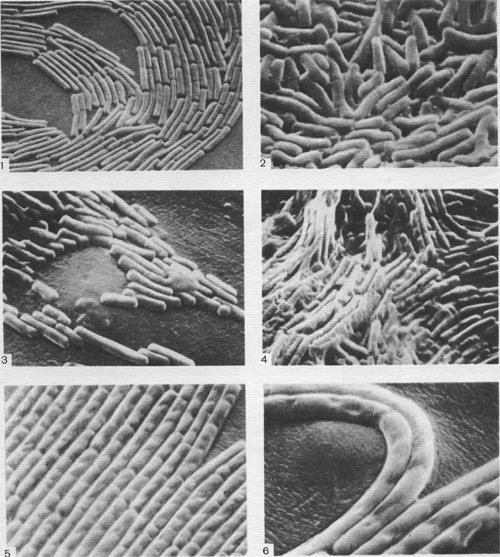
Таблица 37. Расположение клеток в колониях спорообразующих бактерий на поверхности плотных сред: 1—край колонии Bacillus cereus; 2 — центральная часть колонии Вас. cercus; 3 — край колонии Вас. mesentericus; 4 — центральная часть колонии Вас. mesentericus; 5, 6 — тяжи из клеток, образуемые Вас. mycoides. Фотографии сделаны Э. Г. Африкяном в сканирующем электронном микроскопе.
Таким образом, при дыхании конечным акцептором водорода является кислород. У анаэробов в качестве акцепторов водорода выступают либо органические субстраты (брожение), либо неорганические вещества, такие, как нитраты или сульфаты («анаэробное дыхание»). Из схемы видно, что наиболее просто и примитивно транспорт электронов осуществляется у большинства анаэробов из-за отсутствия у них ферментов цепи переноса электронов, способных передавать электроны по цепочке вплоть до молекулярного кислорода.
Молекулярный кислород отрицательно действует на рост и активность облигатных анаэробов. В присутствии свободного кислорода клетки анаэробов теряют подвижность. Благодаря этой реакции анаэробы были впервые обнаружены Пастером. Однажды он рассматривал под микроскопом каплю бродящей жидкости (при маслянокислом брожении), помещенную между двумя тонкими плоскими стеклышками, и заметил, что клетки, находящиеся по краям препарата (куда кислород воздуха . свободно проникал), становились внезапно неподвижными, а палочки, находящиеся в центре препарата (куда воздух не проникал), продолжали очень активно двигаться.
Отсюда Пастер сделал вывод, что кислород воздуха ядовит для некоторых микробов, и разделил последние на две группы — аэробов и анаэробов.
Противники Пастера (например, Трекюль) возражали против утверждения о существовании бактерий, для которых кислород воздуха может быть смертельным, и приводили в пример споры анаэробов, способные длительное время сохраняться на воздухе. На это Пастер отвечал, что споры не являются настоящими живыми существами, так как они не питаются и не размножаются. Последующее развитие науки подтвердило положения Пастера. Так, было показано, что спорам бактерий свойствен крайне выраженный анабиоз и обмен веществ у них находится на таком низком уровне, что его даже не удается измерить с необходимой точностью. В связи с этим споры не чувствительны ко многим повреждающим факторам, а споры анаэробов могут, кроме того (в отличие от вегетативных клеток), легко сохраняться на воздухе.
Вопрос, почему кислород является токсичным для анаэробов, еще недостаточно выяснен. Одни исследователи считают, что токсическое действие кислорода связано с образованием в культурах анаэробных микроорганизмов ядовитых концентраций перекиси водорода, образующейся в результате окисления субстрата кислородом воздуха. Причиной накопления перекиси водорода является отсутствие у анаэробов фермента каталазы (разлагающего перекись). Имеются сообщения, что некоторые штаммы клостридиев могут расти и в аэробных условиях, если к среде добавлена каталаза. Но, с другой стороны, очень важным фактором, определяющим развитие анаэробов, являются окислительно-восстановительные условия среды. Они выражаются через окислительно-восстановительный потенциал (ОВП), измеряемый в вольтах (напряжение на электроде, погруженном в испытуемую среду). Окислительно-восстановительные условия можно выразить также через показатель гНз, характеризующий соотношение между На и 02. rH2 в пределах от 0 до 40 характеризует все степени восстановленности или окисленности среды в зависимости от насыщения ее кислородом либо водородом. Подробные исследования условий развития анаэробов показали, что облигатные анаэробы не могут развиваться при rH2 (или ОВП) выше определенного предела. rНз в среде можно измерить либо электрометрическим способом . (с помощью потенциометров), либо с помощью красителей, которые восстанавливаются и обесцвечиваются (или изменяют цвет) при определенных значениях гНз. Так, например, краситель янусгрюн в аэробных условиях при rH = 20 и выше имеет в растворе зеленый цвет, при rН2 в пределах 12—14 — розовый цвет, а при еще более низких значениях rH он обесцвечивается.
А.Р. Прево проделал следующий опыт (схема 2 на табл. 36). Питательную среду (мясо-пептонный агар или желатин) он наливал в пробирки, кипятил 35 мин и оставлял на воздухе двое суток. Сразу после кипячения среда даже у самой поверхности была сильно восстановленной с низким гН2. Через 48 ч вследствие диффузии кислорода в среду показания гШ сильно изменились. С самой поверхности и до глубины 10 мм гШ составлял 20 единиц (янусгрюн — зеленый). Ниже 10 мм гНа был равен 14 (янусгрюн — розовый). На глубине 13 мм -12,5, при 16 мм — 9,2, а при 17 мм — 8, при 18 мм всего 7,4. Оказалось, что строгие анаэробы могут развиваться в средах с rH не выше 14.
Имеются также виды с высокой степенью анаэробности, которые начинают размножаться лишь при крайне низких значениях rH, приближающихся или почти равных нулю. В области rН2 выше 14 находится зона развития аэробов. В промежуточной области (между аэробиозом и строгим анаэробиозом) могут развиваться микроаэрофильные формы, приспособившиеся к развитию в средах с низкими концентрациями свободного кислорода.
При изучении действия кислорода на раз-. витие облигатных анаэробов было показано, что кислород не оказывает губительного действия на анаэробов, если ОВП среды низкий. Действительно, если к среде добавить восстанавливающие агенты, снижающие ОВП, то некоторые анаэробные микроорганизмы способны развиваться на таких средах в аэробных условиях. В целом анаэробы можно отнести к таким микроорганизмам, рост и развитие которых приурочены к природным субстратам, лишенным свободного кислорода и обладающим низким окислительно-восстановительным потенциалом.
Токсическое действие кислорода воздуха на рост и развитие облигатных анаэробов и тяготение к низкому окислительно-восстановительному потенциалу, по современным представлениям, можно объяснить тем, что молекулярный кислород и высокий ОВП могут служить причиной необратимого окисления жизненно важных ферментов, обусловливающих основные процессы их метаболизма.
Большинство анаэробных спорообразующих бактерий лишено также и других геминовых ферментов. Так, у бактерий из порядков Clostridiales и Plectridiales (по Прево) не обнаружены до настоящего времени цитохромы. Для переноса водорода в ряду субстрат — субстрат ими используются флавиновые ферменты, обладающие свойством аутооксидабильности (самоокисление кислородом воздуха). Хотя ферменты облигатных анаэробов (например, флавиновые) окисляются кислородом, он у них не может служить в качестве физиологического акцептора водорода. Реакция флавиновых ферментов с кислородом направлена на детоксикацию молекулярного кислорода.
Из спорообразующих анаэробов цитохромы (цитохромы бис) обнаружены только у суль-фатредуцирующих микроорганизмов рода De-sulfotomaculum. Эти бактерии также очень чувствительны к кислороду. Таким образом, различия между облигатными анаэробами и аэробами касаются прежде всего ферментативного обеспечения терминального окисления. У анаэробов свободный кислород не может быть использован в качестве конечного акцептора водорода.
В клетках спорообразующих анаэробов в энергетическом обмене веществ играют большую роль флавиновые ферменты.
Не удивительно поэтому, что содержание флавиновых дегидрогеназ в клетках анаэробов (маслянокислые бактерии) значительно выше, чем у аэробов.
Облигатные анаэробы представляют собой, очевидно, пример ранних анаэробных форм жизни. Это согласуется с теорией происхождения жизни на Земле, по которой первичными организмами нашей планеты были анаэробы. Сравнительный биохимический анализ приводит к выводу, что в основе энергетического обмена всех без исключения организмов лежат одни и те же поразительно сходные между собой цепи реакций, не связанных с потреблением свободного кислорода, — реакции, которые происходят в клетках современных анаэробов (по А. И. Опарину).
В связи с этим большое значение имеет также вопрос о локализации в клетках анаэробных микроорганизмов дегидрогеназ. Считалось, что у аэробов дегидрогеназы связаны с мембранами, а у анаэробов, не обладающих цитохрома-ми и окислительным фосфорилированием (кло-стридии), эти ферменты не связаны с мембранами и находятся в цитоплазме только в растворимой форме. Эта точка зрения как будто находится в согласии с данными о том, что ферменты гликолитического цикла (ферменты, участвующие в распаде углеводов в анаэробных условиях) находятся в клетках животных в растворимой форме.
Таким образом создавалось впечатление, что механизмы, обеспечивающие анаэробиоз, в структурном отношении организованы более примитивно. Свидетельством этого является отсутствие в данном случае связи дегидрогеназ с мембранами. Однако недавно было показано, что ферменты гликолитического цикла (в том числе дегидрогеназы) локализуются в плазматической мембране.
Ошибочность прежних данных объяснялась грубостью методов выделения мембран для биохимических анализов. Выделение чистых фракций мембран включает жесткие приемы разрушения клеток и последующее разделение осколков клеток на фракции различных клеточных структур. В процессе таких обработок ферменты, связанные с мембранами, могут легко от них отделиться. Как отмечают известные биохимики Д. Грин и Р. Гольдбергер (1968), именно эта хрупкость мембран вводила в заблуждение два поколения биохимиков, которые считали, что гликолити-ческие ферменты не связаны с мембранами.
Применение электронно-микроскопических методов, позволяющих изучать распределение дегидрогеназ в целых клетках, показывает, что дегидрогеназы у анаэробных спороносных бактерий, очевидно, также связаны с мембранами, играющими у живых организмов огромную роль, особенно в процессах энергетического метаболизма. Например, у такого облигатного анаэроба, как Cl. taeniosporum, активность окислительно-восстановительных ферментов проявлялась только в связи с цитоплазматиче-ской мембраной (рис. 60). В то же время у других анаэробов восстановление акцепторов электронов наблюдается и в цитоплазме. Возможно, эти явления связаны с различным набором ферментов у разных видов либо с неспецифическим восстановлением красителей в цитоплазме.
В качестве индикаторов для окислительно-восстановительных ферментов в электронной микроскопии используют теллурит калия или соли тетразолия. Эти соединения, будучи добавленными в среду и проникая в клетки, акцептируют электроны, которые они получают от дегидрогеназ. В результате реакции эти соединения восстанавливаются и выпадают в осадок, имеющий вид электронно-плотных (темных на экране микроскопа) зернышек, глыбок, тонких слоев. Теллурит в качестве индикатора окислительно-восстановительных ферментов более показателен, чем тетразолии, так как соли тетразолия могут неспецифически восстанавливаться веществами, содержащими сульфгид-рильные группы, а также редуцирующими (восстанавливающими) сахарами. Особенно это относится к анаэробным микроорганизмам, развивающимся при очень низком окислительно-восстановительном потенциале среды и вырабатывающим различные вещества с сильной редуцирующей активностью. Весьма интересно, что у некоторых анаэробных бактерий рода Clostridium обнаружена специфическая, но эффективная форма окислительного фосфорилиро-вания (фосфорилирования в цепи окислительных ферментов).
Если согласиться с утверждением (довольно убедительно обоснованным данными сравнительной физиологии и биохимии) о том, что облигатные анаэробы — это пример ранних форм жизни на Земле, то возникает вопрос, не отразились ли происхождение и эволюция анаэробов на составе и строении их ДНК — хранителе генетической информации. Сейчас уже хорошо известно, что дезоксирибонуклеиновым кислотам всего органического мира присущ единый план строения, а с другой стороны, имеются безграничные возможности вариаций состава и структуры этих соединений. Вполне логично думать, что возникновение ДНК в истории жизни на Земле было весьма важным и, вероятно, даже решающим фактором в дифференциации и обособлении новых групп и видов живых существ. Так как именно нуклеиновые кислоты непосредственно связаны с наследственностью и изменчивостью, то они должны являться материальной основой эволюционного процесса.
Специфичность нуклеиновых кислот для различных организмов обусловливается их химическим строением и может отражаться в первую очередь на количественном соотношении пуриновых и пиримидиновых оснований (на нуклеотидном составе) и на последовательности чередования нуклеотидов в цепи молекулы. Состав ДНК бактерий очень сильно варьирует от самого крайнего АТ-типа (преобладает пара — аденин и тимин) до ГЦ-типа (преобладают гуанин и цитозин). Если выразить соотношение оснований в виде молярных процентов Г + Ц, то колебания этого показателя лежат в пределах от 25 до 75%. Весьма показательно в этом отношении, что сложно организованные прокариотные организмы -актиномицеты — имеют самый высокий процент Г + Ц в ДНК, а примитивные анаэробные спорообразующие бактерии, у которых отсутствуют цитохромы, обладают крайним АТ-типом(до25% Г + Ц).Средиспорообразующих анаэробов также можно выявить определенные закономерности. Так, для гетеротрофных, не содержащих цитохромов бактерий (клостридии) колебания Г + Ц находятся в пределах от 25 до 39%, а для спорообразующих сульфатредуцирующих анаэробов, являющихся хемолито-гетеротрофами и обладающих уже отдельными цитохромами, процент Г + Ц находится в пределах 45—48.
Таким образом, ДНК у анаэробов имеет интересные особенности, и очень многого можно ожидать в будущем от изучения первичной структуры ДНК этих бактерий.
Дата добавления: 2015-06-12; просмотров: 1533;