Спиртового брожения
Основные возбудители спиртового брожения - дрожжи - традиционно используются в хлебопечении, виноделии, получении спирта, пивоварении. По значимости с ними конкурируют только молочнокислые бактерии.
При производстве рисового пива (саке) в Японии в качестве инициатора спиртового брожения используют Aspergillus orysae. Спиртовое брожение могут вызывать бактерии - Zymomonas mobilis, Z.anaerobica, Sarcina ventricula, Erwinia amylovora. Но получение винного спирта с помощью бактерий промышленного значения не приобрело.
Дрожжи относятся к грибам, но в систематике грибов отдельного единого таксона не составляют, термин "дрожжи" таксономического значения не имеет. В настоящее время известно более 500 видов. В зависимости от типа полового процесса и его наличия распределены по трем классам: Ascomycetes, Basidiomycetes, Deuteromycetes (Fungi imperfecti). Аскомицетовые дрожжи объединены в порядок Endomycetales. Они характеризуются примитивным половым процессом (слияние двух вегетативных клеток), не формируют плодовые тела (протоаскомицеты), размножаются почкованием или делением. Зигота, образующаяся в результате полового процесса, превращается в аск с эндогенными половыми спорами. В аске содержится от одной до четырех, реже восьми спор. Только у дрожжей родов Lipomyces и Kluyveromyces некоторые виды в аске имеют до нескольких десятков спор. К аскомицетам относятся широко применяемые в качестве продуцентов этанола дрожжи родов Saccharomyces и Schizosaccharomyces. Базидиальные дрожжи образуют половые структуры базидиального типа (экзогенные половые базидиоспоры). К ним относятся пигментированные (красные) дрожжи рода Rhodosporidium, непигментированные - Leucosporidium, дрожжи рода Sporobolomyces, размножающиеся баллистоспорами.
К несовершенным дрожжам (Deuteromycetes) относят дрожжи и дрожжеподобные грибы, не имеющие полового цикла, образующие только бесполые споры (бласто-, артро-, хламидоспоры), размножающиеся преимущественно почкованием. Организмы, принадлежащие к этой группе, произошли от высших грибов в процессе утраты ими половых функций. Наиболее распространенными продуцентами белка в микробиологической промышленности являются несовершенные дрожжеподобные грибы рода Candida.
Дрожжи растут на поверхности сладких плодов, в нектаре цветков, в сокотечениях деревьев, на поверхности листьев, в лесном и травяном опаде и почве. Их можно встретить в водоемах, в пищеварительном тракте человека и животных. Большинство дрожжей сапрофиты, но встречаются патогенные (болезнетворные) и условнопатогенные формы.
Из соединений углерода дрожжи лучше всего используют гексозы, а также некоторые виды пентоз. Из полисахаридов чаще всего утилизируют инулин и крахмал. Встречаются виды, растущие на средах с углеводородами, спиртами (в т.ч. метанолом и этанолом), органическими кислотами и другими углеродными субстратами.
Источником азота дрожжевым клеткам служат соли аммония, аминокислоты, пептиды, реже нитраты и нитриты. Некоторые виды нуждаются в факторах роста - витаминах, чаще в биотине и тимине, другие - способны все витамины синтезировать сами.
Границы рН сред, при которых растут дрожжи, от 3,0 до 8,0 ед., оптимальные значения рН от 3,5 до 6,5 ед. Температурный диапазон для роста достаточно широк: от 00С (в некоторых случаях - минус 70С) до 48…500С, но оптимальные температурные значения для большинства видов 28…300С, хотя расы дрожжей, используемых в пивоварении, имеют более низкий оптимизм. Облигатно психрофильные формы не растут при температуре выше 18…200С. Большинство дрожжей - факультативные анаэробы. В анаэробных условиях получают энергию в результате сбраживания углеводов, в аэробных - за счет дыхания. В отличие от гликолиза у высших организмов, на последних этапах которого образуется молочная кислота, у дрожжей образуется этиловый спирт. Такой процесс возможен из-за наличия фермента пируватдекарбоксилазы, катализирующей переход пирувата в ацетальдегид, с последующим восстановлением его в этанол.
Гликолитическим (Эмбдена-Мейергофа-Парнаса) или его еще называют фруктозобисфосфатным (ФБФ) путем осуществляется разложение глюкозы, галактозы, фруктозы, маннозы. Олигосахариды должны вначале гидролизоваться ферментами до гексоз.
Существовало мнение, что только в аэробных условиях пентозы могут усваиваться дрожжами. Сравнительно недавно установлена способность некоторых видов к росту на средах, содержащих ксилозу или ксилулозу, в анаэробных условиях с образованием этанола. Это очень важно для производств спирта на основе гидролизных сахаров, полученных при переработке древесины и отходов сельскохозяйственных растений. Разложение высших спиртов и пентоз дрожжами осуществляется через пентозофосфатный и фруктозобисфосфатный пути, при этом спирты сначала дегидрируются до гексоз и пентоз.
Дрожжи родов Rhodotorula, Sporobolomyces, Cryptococcus метаболизируют сахара только в аэробных условиях, это связано с отсутствием пируватдекарбоксилазы и/или НАД-зависимой алкогольдегидрогеназы - ферментов, завершающих переход пирувата в этанол.

При брожении в этом случае пируват теряет карбоксильную группу под действием пируватдекарбоксилазы, но реального окисления пирувата не происходит. В клетке реакция необратима. Для проявления каталитической активности пируватдекарбоксилазе требуется Mg2+. С молекулой этого фермента прочно связан кофермент тиаминпирофосфат, функция которого декарбоксилирование a-кетокислоты, перенос ацетальдегидных групп. На последнем этапе ацетальдегид восстанавливается до этанола за счет NADH, образовавшегося при окислении глицеральдегид 3-фосфата, реакция катализируется алкогольдегидрогеназой. Суммарное уравнение спиртового брожения имеет вид:
Глюкоза + 2ФН + 2АДФ ® 2 Этанол + 2СО2 + 2АТФ + 2Н2О
Нейберг назвал этот процесс первой формой брожения.
В зависимости от условий культивирования процесс брожения может изменяться. При включении в питательную среду бисульфита натрия происходит связывание ацетальдегида, который блокируется и исключается из процесса.

В этом случае акцептором электронов (Н+) от NADH становится дигидроксиацетонфосфат, трансформирующийся в глицерин-3-фосфат, а затем в глицерин. Это вторая форма брожения по Нейбергу, суммарная реакция которой выражается следующим уравнением:
Глюкоза ® глицерин + ацетальдегид (связанный) + СО2
Суммарное количество синтезированной АТФ при таком варианте брожения равно 0, поэтому обеспечить рост клеток процесс не может, однако, он используется для промышленного получения глицерина.
Очень сходен вариант спиртового брожения при культивировании дрожжей в щелочной среде. В этом случае ацетальдегид окисляется НАД-зивисимой дегидрогеназой в уксусную кислоту. Образовавшийся НАДН используется для восстановления эквивалентного количества ацетальдегида в этанол НАДН, получающийся при окислении 3-фосфоглицеринового альдегида, восстанавливает дегидроксиацетонфосфат в глицерин-3-фосфат, который превращается в глицерин. Это третья форма брожения по Нейбергу, выражается уравнением:
2Глюкоза+Н2О®2глицерин + этанол + уксусная кислота + 2СО2
Такое направление процесса благоприятно для клеток, образующаяся уксусная кислота снижает рН среды, после этого возобновляется нормальное спиртовое брожение.
 В начальной стадии спиртового брожения дегидроксиацетонфосфат выполняет роль акцептора электронов до накопления ацетальдегида, необходимого для окисления НАДН, на это указывает существование периода индукции в начале брожения, при котором появляется глицерин.
В начальной стадии спиртового брожения дегидроксиацетонфосфат выполняет роль акцептора электронов до накопления ацетальдегида, необходимого для окисления НАДН, на это указывает существование периода индукции в начале брожения, при котором появляется глицерин.
Рисунок 5 - Схема глицеринпировиноградного брожения
Одновременно 3-фосфоглицериновый альдегид превращается согласно фруктозобисфосфатному пути в пируват, который декарбоксилируется в ацетальдегид, а он восстанавливаться в спирт не может, так как для образования глицерина из дегидроксиацетонфосфата использован весь НАДН. При образовании в процессе брожения молекулы глицерина накапливается одна молекула пировиноградной кислоты или ацетальдегида. Превращения в спирт не происходит. Спиртовое и глицеринпировиноградное брожение тесно связаны. Вначале преобладает глицеринпировиноградное брожение, но наряду со спиртом обнаруживаются и другие продукты метаболизма. В этом случае необходимые для биосинтеза кислоты ЦТК образуются карбоксилированием пирувата:
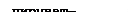
Пируват + АТФ + СО2 + Н2О Оксалоацетат + АДФ + ФН
На первых этапах спиртового брожения преобладает брожение глицеринпировиноградное, при этом образуется глицерин и пируват. Пировиноградная кислота идет на формирование вторичных продуктов: уксусной, молочной, янтарной, пропионовой, муравьиной и других кислот, ацетона, диацетила, ацетоина, 2,3 бутандиола, альдегидов и сложных эфиров. В процессе сбраживания сахаров дрожжами накапливается незначительное количество D(-)-молочной кислоты, однако, Sacch.veronae синтезирует L(+)-молочную кислоту. Из пирувата в небольшом количестве образуются лимонно-яблочная и диметилглицериновая кислоты.
При спиртовом брожении образуются побочные продукты - высшие спирты (сивушные масла): изоамиловые (3-метилбутанол и 2-метилбутанол), изобутиленовый, н-бутиловый, н-пропиловый, ароматические (b-фенилэтиловый, н-оксифенилэтиловый) спирты. Эти продукты синтезируются из кето- или аминокислот. Они влияют на вкус и аромат конечного продукта. Содержание их зависит от исходного субстрата, включающего сбраживаемые углеводы, и не коррелирует с образованием этанола, сброженные растворы, содержащие одинаковые количества спирта, могут значительно отличаться по вкусовым и ароматическим качествам. Процесс образования высших спиртов включает дезаминирование амино- в кетокислоту, которая декарбоксилируется в альдегид, восстанавливаемый в спирт.
В присутствии О2 дрожжи быстро переходят на аэробное дыхание и пируват через цикл трикарбоновых кислот (ЦТК) окисляется до СО2 и Н2О. В процессе ЦТК клетка обеспечивается метаболитами, необходимыми для последующих биосинтетических реакций. В энергетическом плане дыхание более выгодно, чем брожение, дрожжи при этом растут активнее, образуя большую биомассу.
Подавление брожения в аэробных условиях носит название эффекта Пастера. Он связан с различием энергетического заряда клеток в аэробных и анаэробных условиях. Между дыхательной системой и субстратным фосфорилированием происходит конкуренция за АДФ. Значение имеет и аллостерическая регуляция фосфофруктокиназы. Фермент ингибируется АТФ и активируется АМФ. В анаэробных условиях содержание АТФ низкое, но активность фермента высока. В аэробных - отношение АТФ/АМФ возрастает, активность же фосфофруктокиназы снижается. Аллостерическим ингибитором фосфофруктокиназы является цитрат - промежуточный метаболит в цикле трикарбоновых кислот.
Спиртовое брожение может происходить в условиях повышенной аэрации при содержании глюкозы в среде от 1,5 до 2,0%. Этот эффект при высокой скорости усвоения глюкозы, наблюдаемый в данном случае, называется катаболической репрессией или эффектом Крэбтри. Он не отмечается при выращивании дрожжей на питательных средах, включающих слабо усваиваемые сахара. Катаболитная репрессия аэробного дыхания сопровождается снижением получения клетками энергии, подавлением биосинтеза промежуточных продуктов цикла трикарбоновых кислот и глиоксилатного цикла.

Не все высшие спирты синтезируются из аминокислот. Другой путь - биосинтез из продуктов метаболизма углеводов. При конденсации пировиноградной с уксусным альдегидом или ацетил-КоА образуется ацетомолочная кислота, которая трансформируется в 2-кетоизовалериановую кислоту, в результате ее декарбоксилирования получается изомасляный альдегид, восстанавливающийся в изобутанол.

При спиртовом брожении образуются и серосодержащие вещества сероводород, сульфиты, меркаптаны с неприятным запахом. Выделение СО2 при брожении способствует удалению этих веществ из культуральной среды.
Дата добавления: 2015-06-12; просмотров: 2911;