Видообразование и половой отбор

В истории науки бывали случаи, когда хорошая идея, давно уже высказанная каким-нибудь гениальным теоретиком и даже подкрепленная фактами, не находила всеобщего признания до тех пор, пока кто-нибудь не разрабатывал математическую модель, которая объяснила бы всем, как и почему эта идея работает. Больше всех не повезло теории полового отбора. Эта абсолютно гениальная (как мы теперь понимаем) идея была разработана Дарвином, но современники ее не поняли и не приняли. Современники Дарвина готовы были признать, что самцы конкурируют за самок, и этим можно объяснить развитие «вооружения» вроде оленьих рогов. Но что самки могут активно выбирать самцов – в это они не могли поверить. Слишком безнравственно. Самки, по мнению мыслителей Викторианской эпохи, могут лишь скромно сидеть в уголочке в нарядном платье и ждать, пока к ним кто-то посватается. А без активного выбора самками нельзя объяснить такие вещи, как павлиний хвост (украшения).

Идея полового отбора успела стать «полузабытым научным курьезом», когда в 1930 году ее эксгумировал ее, развил и дополнил важными деталями Рональд Фишер. ( - один из создателей мат. статистики и генетической теории эволюции; странности, евгеника).

Что произошло? Ведь самок никто не отбирал, только самцов!
Именно Рональд Фишер первым понял, что не только брачные украшения самцов, но и вкусы и избирательность самок тоже наследуются и эволюционируют – по тем же правилам, что и остальные признаки (Дарвин предполагал, что это какое-то врожденное чувство прекрасного).
Эта простая мысль сделала теорию полового отбора полностью работоспособной. Однако идеи Фишера о половом отборе тоже не были восприняты научным сообществом. Они почти забылись к тому времени, когда их снова выкопали из забвения представители следующего поколения биологов-теоретиков, сумевшие, наконец, разработать убедительные и понятные математические модели. Лишь с третьей попытки теория полового отбора получила заслуженное признание.

Три важнейших идеи (модели), развитые в рамках теории полового отбора: фишеровское убегание, индикаторы приспособленности, принцип гандикапа.
ФИШЕРОВСКОЕ УБЕГАНИЕ.
· Возможность взрывообразной, «автокаталитической», согласованной эволюции какого-нибудь произвольного признака и «моды» на этот признак среди особей противоположного пола.
· Сама мода на признак делает его адаптивно выгодным (полезным). Полезность признака, в свою очередь, делает адаптивно выгодной «моду» на него. Получается положительная обратная связь: чем большему числу самок нравятся самцы с этим признаком, тем выгоднее самцам иметь этот признак, а чем выгоднее признак, тем выгоднее для самок строго придерживаться этой моды.

Допустим, в популяции появился мутантный аллель, влияющий на брачные предпочтения самок. Допустим, самки с этим геном выбирают самых длинноухих самцов. В популяции существует небольшая нейтральная (не влияющая на приспособленность) изменчивость по длине ушей. Самки с мутантным геном поначалу не имеют никакого преимущества, но и особого вреда их странные вкусы им не приносят. Поэтому мутантный аллель имеет шанс за счет дрейфа достичь некоторой заметной частоты в генофонде популяции. Вот тут‑то и вступает в действие механизм “убегания”. Длинноухие самцы получают репродуктивное преимущество, потому что им доступны все самки, а короткоухие самцы могут спариться только с теми самками, у которых нет мутантного аллеля. Длинноухие самцы начинают оставлять больше потомков, чем короткоухие.
Потомство от браков длинноухих самцов с мутантными самками наследует не только “гены длинноухости” (от отца), но и аллель предпочтения длинноухих самцов (от матери). В результате эти аллели начинают распространяться.
Как только длинноухие самцы начинают оставлять в среднем больше потомства, чем короткоухие, самкам становится выгодно выбирать длинноухих партнеров, потому что тогда их сыновья унаследуют длинноухость, привлекут больше самок и оставят больше потомства. В результате те самки, которые предпочитают длинноухих самцов, начинают оставлять больше внуков, то есть получают репродуктивное преимущество.
Бывшая случайная прихоть превращается в полезную адаптацию. Возникает положительная обратная связь, в результате которой в генофонде быстро распространяются гены длинноухости и гены любви к длинноухим.
Длинноухость выгодна, потому что самки любят длинноухих, а любить длинноухих выгодно, потому что выгодна длинноухость. И никому нет дела до того, нужны ли длинные уши зачем‑то еще. В этом и состоит суть фишеровского убегания.
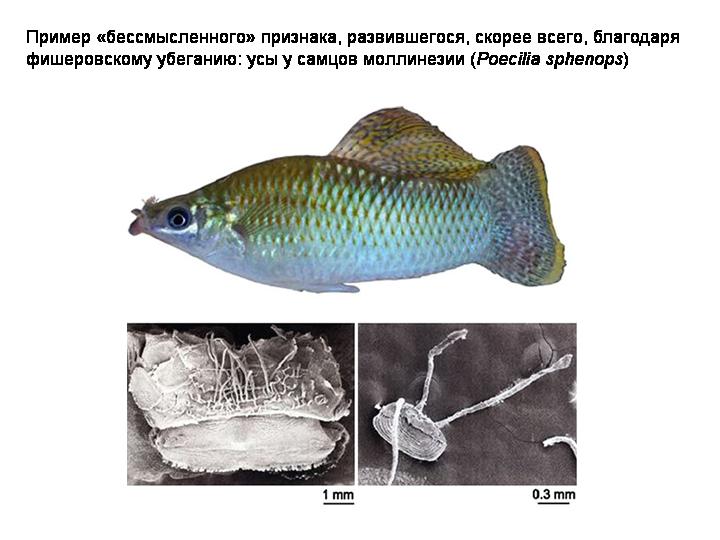
Пример «бессмысленного» признака, развившегося, скорее всего, благодаря фишеровскому убеганию.
ИНДИКАТОРЫ ПРИСПОСОБЛЕННОСТИ
· Самкам выгодно выбирать самцов по признакам, свидетельствующим о хорошем здоровье, а значит, о «качественных» генах.
· Поэтому в большинстве случаев фишеровское убегание подхватывает не бессмысленные признаки, а «индикаторы приспособленности».

· Если «индикатор приспособленности» обходится самцу слишком дешево, обязательно расплодятся «обманщики», и выбирать по этому признаку станет невыгодно.
· Индикаторы приспособленности (под действием убегания) часто становятся обременительными, так что слабый самец не может их имитировать (они ему «не по карману»).
· Обременительные и.п. более эволюционно стабильны, чем дешевые. Дорогая реклама и «дешевая болтовня».

Половой отбор — мощная эволюционная сила, способная многократно усиливать эффективность «обычного» естественного отбора и ускорять адаптивную эволюцию (а не только создавать бессмысленные украшения и причудливое брачное поведение). Если самки выбирают самцов не по произвольным бессмысленным критериям, а по признакам, отражающим приспособленность или «качество генов» самца, то тем самым самки резко ускоряют адаптивную эволюцию. Половой отбор в этом случае становится мощным усилителем обычного, естественного отбора.
Каким образом половой отбор повышает эффективность естественного отбора? Самец с пониженной приспособленностью не только имеет меньше шансов выжить, но и становится менее привлекательным для самок. Ситуация «мало того, что здоровье слабое, так еще и девушки не любят». Даже очень небольшие различия в приспособленности, едва заметные для обычного отбора, могут стать решающими, когда дело доходит до конкуренции между самцами в попытках очаровать привередливую самку.

Кроме того, отбор перестает быть «слепым». Теперь его направляют существа, у которых всё же есть кое-какие мозги. Эти существа способны к целенаправленным действиям и отчасти осмысленным решениям. Самки становятся селекционерами. Привередливые павлинихи создали роскошные узоры на хвостах самцов-павлинов точно так же, как голубеводы создали причудливые украшения у декоративных пород голубей.
П.О. СПОСОБСТВУЕТ СИМПАТРИЧЕСКОМУ ВИДООБРАЗОВАНИЮ
Половой отбор может способствовать симпатрическое видообразование. Ясно, что без полового отбора и избирательного скрещивания симпатрическое видообразование едва ли возможно. До недавних пор все думали, что для симпатрического видообразования самки двух зарождающихся видов должны иметь разные половые предпочтения. И было не очень понятно, с чего бы вдруг у самок в одной популяции возникли разные вкусы. Но на самом деле, как было недавно показано, половой отбор стимулирует симпатрическое видообразование даже в том случае, если вкусы у всех самок остаются одинаковыми.
Сандер ван Дорн из Бернского университета (Швейцария) и его коллеги в 2009 г опубликовали важную статью, которая должна окончательно превратить симпатрическое видообразование в общепринятую теорию. Размах чувствуется уже в названии статьи. Без ложной скромности авторы озаглавили ее «О происхождении видов путем естественного и полового отбора», что является точным повторением заглавия книги Дарвина, за исключением предпоследнего слова.
Модель ван Дорна показывает, что даже если все самки в популяции имеют одинаковые половые предпочтения, их привередливость может радикально повысить вероятность симпатрического видообразования.
Понятно, что половой отбор по индикаторам приспособленности повышает эффективность обычного отбора – как отрицательного (очищающего), так и положительного отбора, то есть фиксации полезных мутаций.
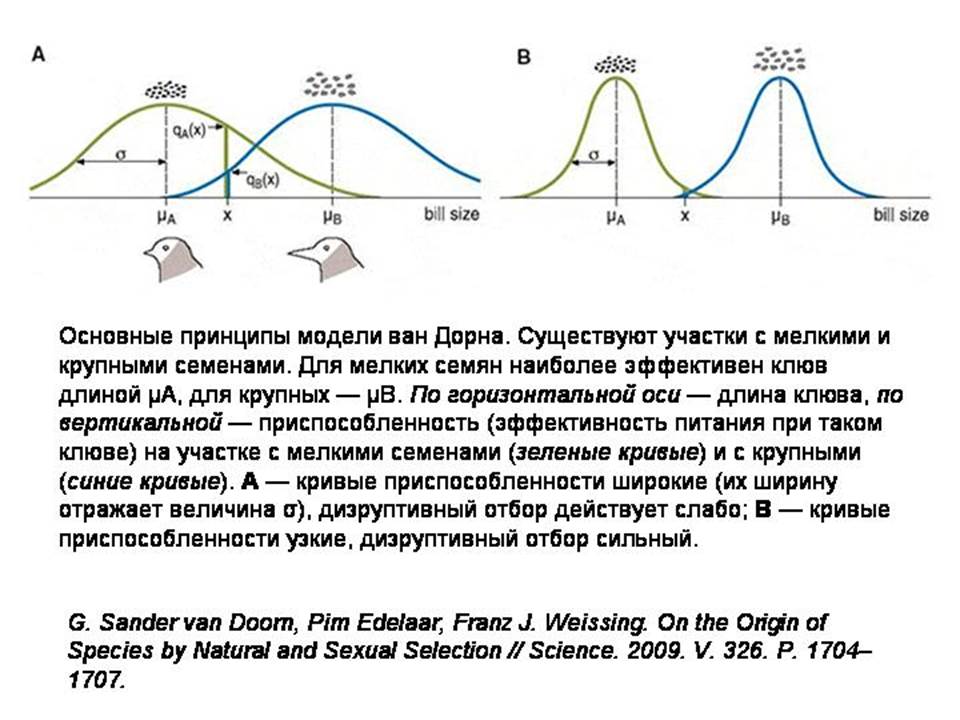
Основная идея ван Дорна и его коллег состоит в том, что отбор по «индикаторам приспособленности» повышает также и эффективность дизруптивного (разрывающего, разводящего) отбора. Дизруптивный отбор действует в том случае, если крайние варианты какого-то признака более адаптивны, чем промежуточные.
Допустим, существует популяция птиц с клювами средней длины. Местность, где они живут, состоит из участков двух типов. На одних участках преобладают растения с крупными семенами, на других — с мелкими. Для разгрызания крупных семян удобнее иметь длинный клюв, для мелких — короткий. В такой ситуации может начаться дизруптивный отбор. Это значит, что преимущество получат птицы с более длинными или более короткими клювами, но не с промежуточными. Известно, что у реальных птиц, попавших в подобные условия, может произойти симпатрическое видообразование. Остается только понять, каким образом видам удается разделиться, если их представители, перелетая с участка на участок, то и дело встречаются друг с другом и никто не мешает им образовывать смешанные пары и производить гибридное потомство с промежуточными по длине клювами.
В модели все птицы изначально имеют одинаковые клювы средней длины, у самцов нет «индикаторов приспособленности», а у самок — избирательности при выборе партнера, поэтому скрещивания происходят случайным образом. Все признаки могут меняться в результате мутаций. Взрослые птицы живут на своих участках, молодые могут мигрировать на другие участки. Специальная переменная задает интенсивность миграций.
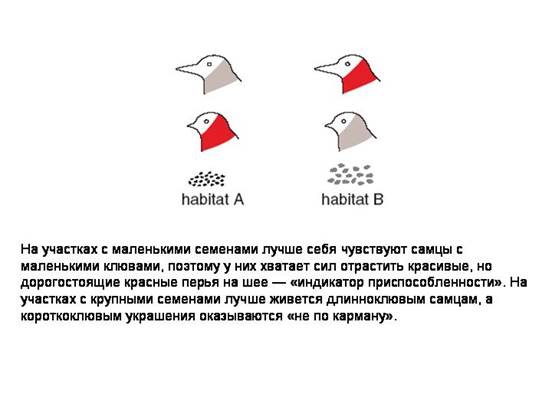
На участках с крупными семенами лучше себя чувствуют особи с большими клювами, и наоборот. Птица, которая лучше питается, имеет больше шансов дожить до зрелости. Кроме того, ее организм располагает большим количеством ресурсов. Самцы могут направить часть этих ресурсов на формирование красного оперения, которое служит «индикатором приспособленности». Самки могут приобрести способность выбирать самцов по этому признаку.
И красное оперение, и избирательность самок — «дорогостоящие» признаки. Это значит, что на их развитие тратится часть ресурсов организма, которые могли быть использованы на повышение выживаемости. Самцы платят за свою красоту снижением шансов на выживание. Именно поэтому данный признак может служить индикатором приспособленности. Если бы красное оперение было «бесплатным», все самцы быстро стали бы профессиональными обманщиками, а избирательность самок потеряла смысл.
Главными параметрами модели являются интенсивность миграций молодых птиц и ширина кривой приспособленности (величина σ на рисунке). Чем больше сигма, тем легче разгрызать семена неподходящим по размеру клювом. Вместе эти две величины задают интенсивность дизруптивного отбора.
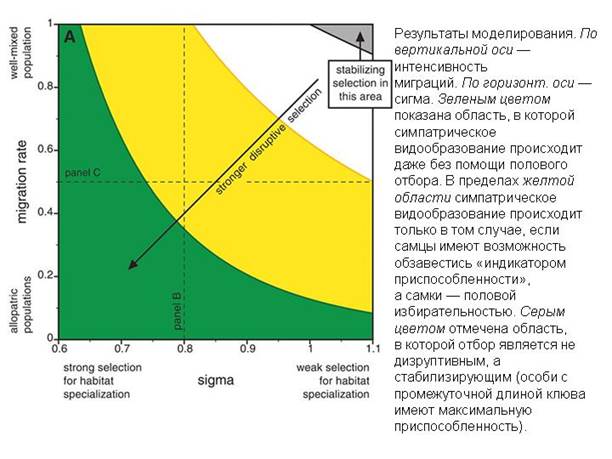
Сначала авторы исследовали поведение модели без «индикатора приспособленности» и половой избирательности. В такой ситуации симпатрическое видообразование происходит только при экстремальных значениях параметров: либо при полном отсутствии миграций, либо при очень маленькой сигме. В остальных случаях силы дизруптивного отбора не хватает, чтобы «разорвать» популяцию надвое.
Затем модельным птицам «разрешили» приобретать в результате мутаций половую избирательность и способность расходовать часть ресурсов на красные перышки. Эффект оказался неожиданно сильным: теперь исходный вид стал распадаться надвое даже при активной миграции и большой сигме. Таким образом, модель показала, что половой отбор по индикатору приспособленности способен радикально повысить вероятность симпатрического видообразования.
РЕЗУЛЬТАТЫ МОДЕЛИРОВАНИЯ.
Рассмотренный в этой модели механизм — не единственный возможный «катализатор» симпатрического видообразования. Симпатрическое видообразование станет еще более вероятным, если «разрешить» птицам в ходе эволюции менять алгоритм миграций. Например, птицы могут выработать алгоритм «мигрируй, только если вокруг нет удобных для тебя семян» или «селись только там, где много удобных семян». Это приведет к снижению частоты гибридизации. Еще одна возможность – выбор партнера «с оглядкой на себя» или с использованием импринтинга (запечатления). Например, самка может предпочитать партнеров с таким же клювом, как у нее самой или у ее отца.
ПРИМЕР ПАРАЛЛЕЛЬНОГО СИМПАТРИЧЕСКОГО ВИДООБРАЗОВАНИЯ НА ОСТРОВАХ ТРИСТАН-ДА-КУНЬЯ
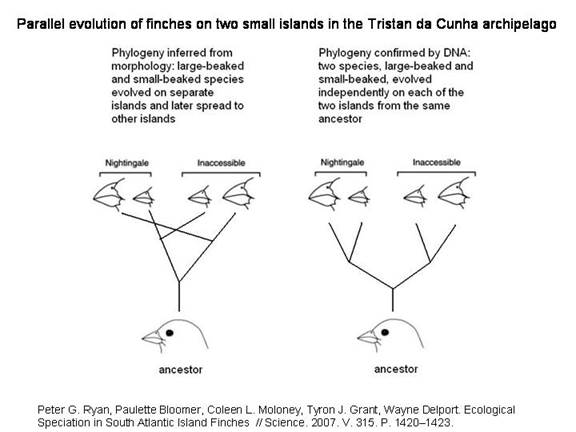
Считалось, что вьюрки рода Nesospiza, обитающие на двух маленьких островах архипелага Тристан-да-Кунья, относятся к двум видам: один с большим клювом, другой с маленьким. Генетический анализ показал, что эти птицы произошли от одной предковой формы, которая когда-то заселила оба острова, а затем на каждом из них подразделилась на большеклювую и мелкоклювую разновидности.
-------------------------------------------


-------------------------------------------------------------------
Дата добавления: 2015-06-10; просмотров: 3467;