БАЛАНСИРУЮЩИЙ ОТБОР
· Такой алгоритм выбора брачного партнера (предпочтение особей с аллелями ГКГ, отличающимися от своих собственных) дает преимущество редким аллелям.
· В результате получается балансирующий («частотно-зависимый») отбор.
· Это ведет к росту и поддержанию высокого уровня полиморфизма аллелей ГКГ в популяции.
· Отбор, осуществляемый вирусными инфекциями, работает в том же направлении.
· Другие примеры балансирующего отбора: соотношение полов, "заботливые отцы" и "проныры".
Часть 5. Биология развития (лекции 14-17)

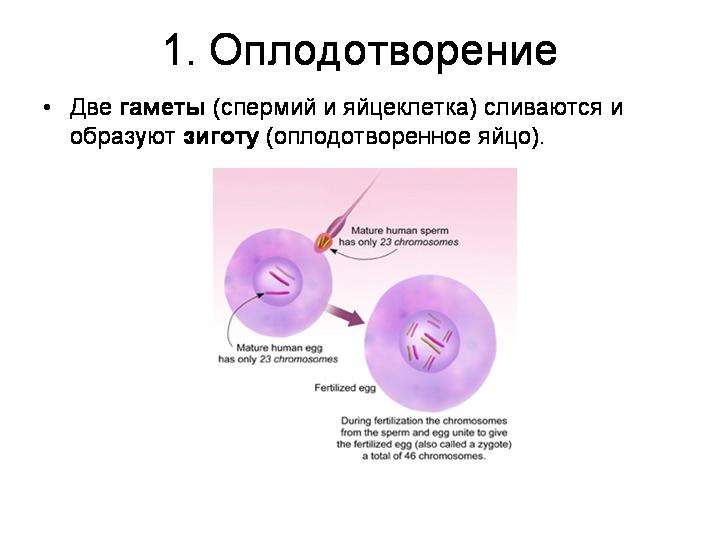
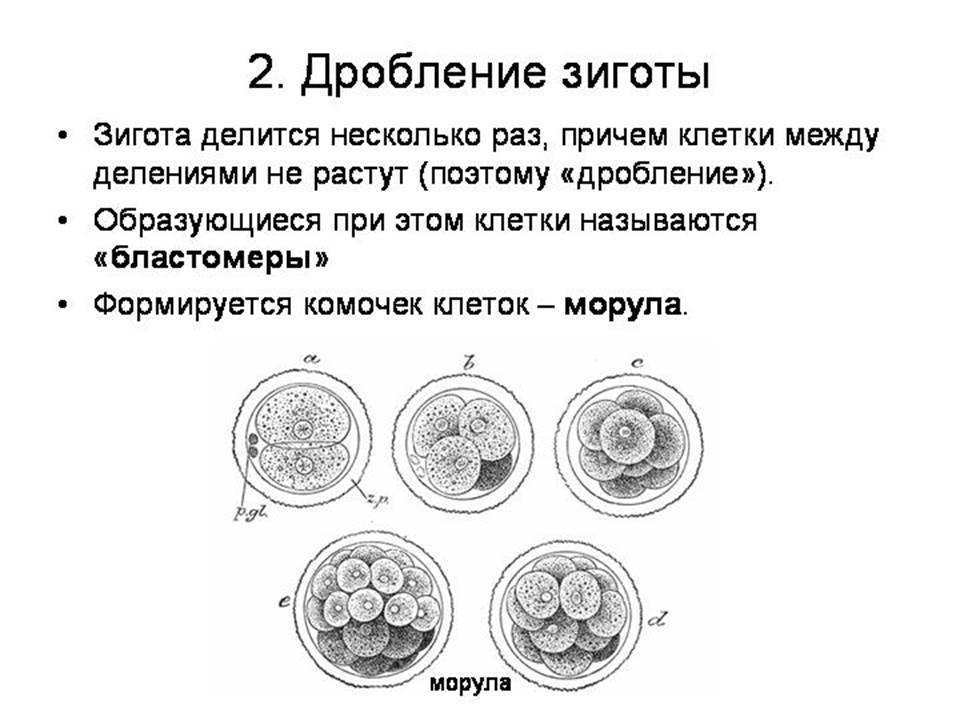
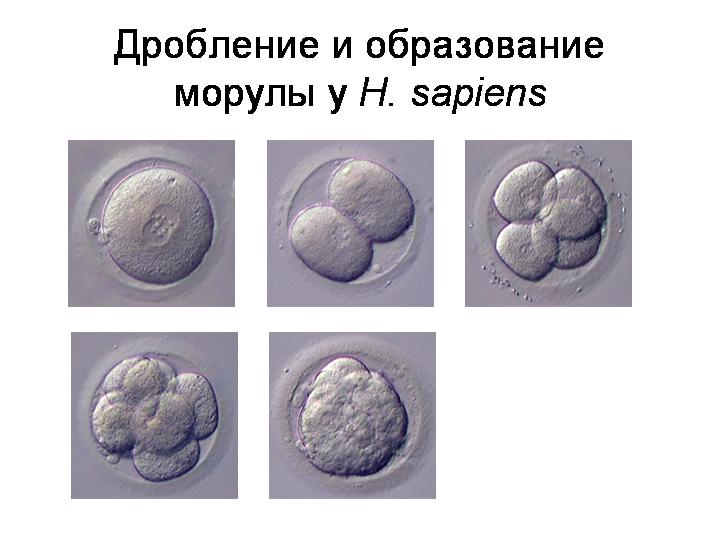
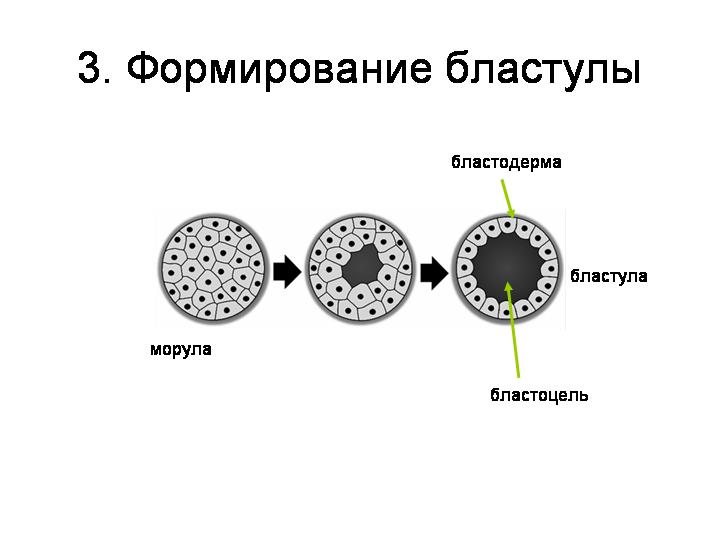
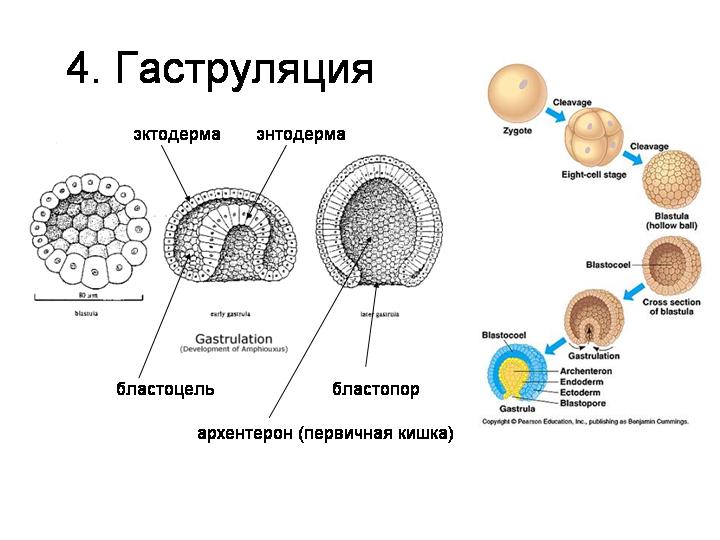
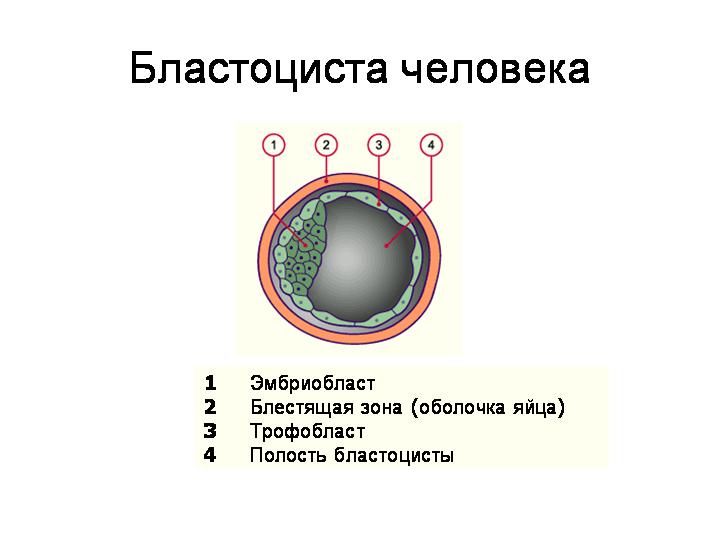
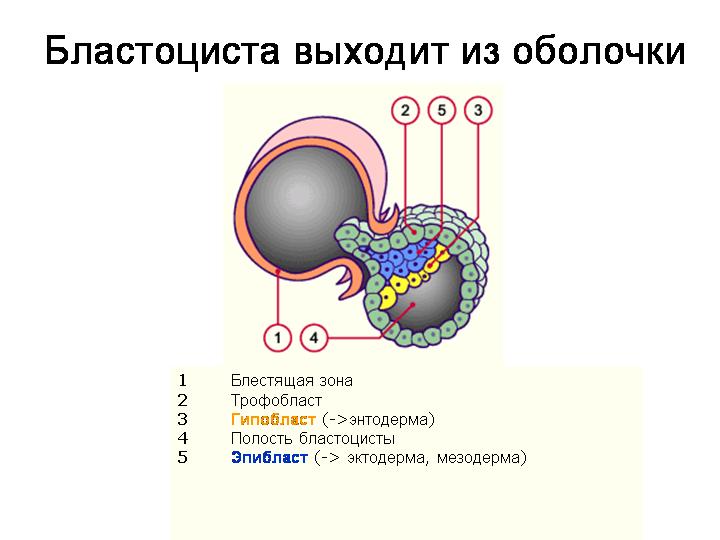
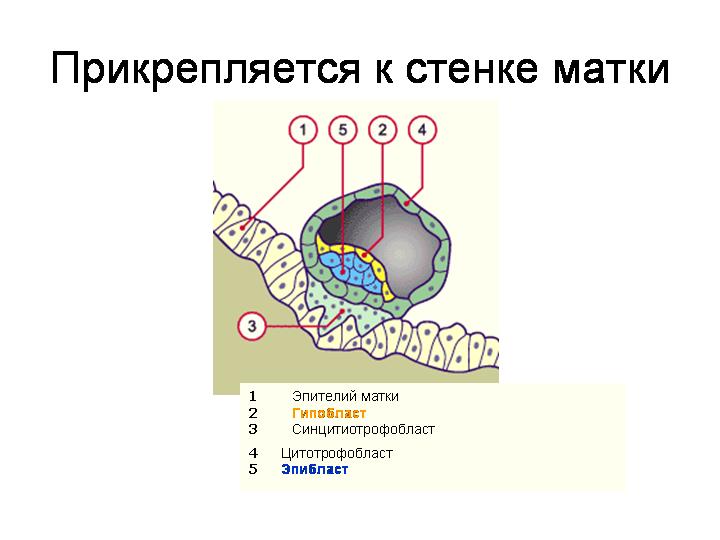
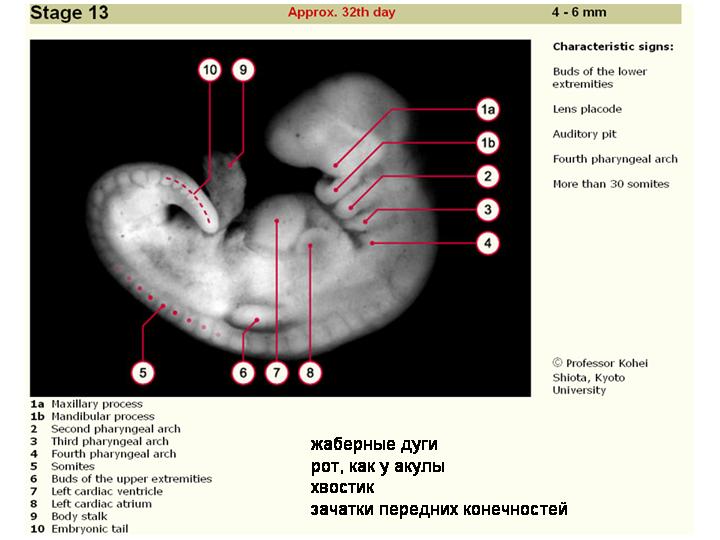
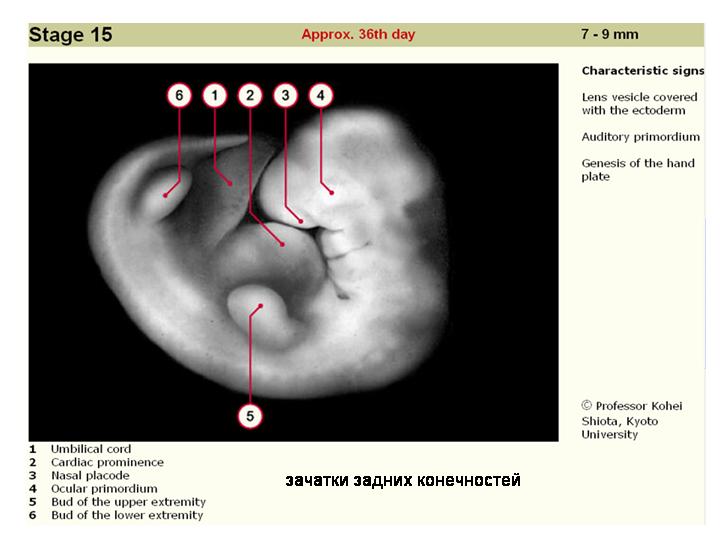
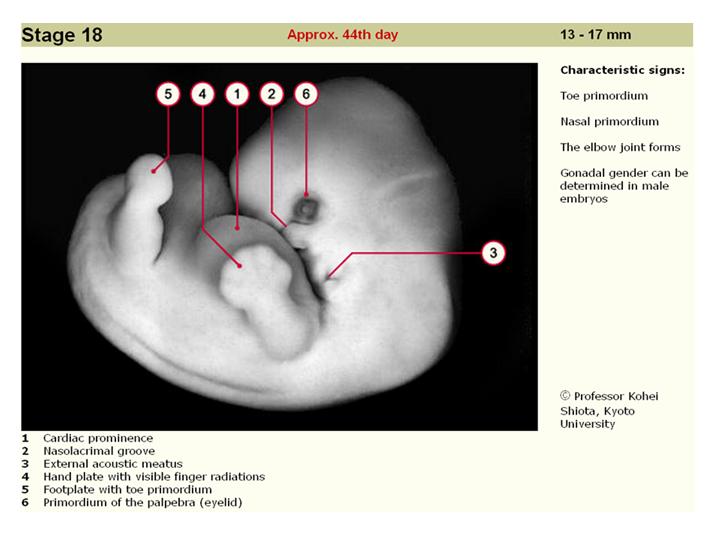
--------------------------------------------------------

ПРИНЦИПИАЛЬНЫЙ ВОПРОС
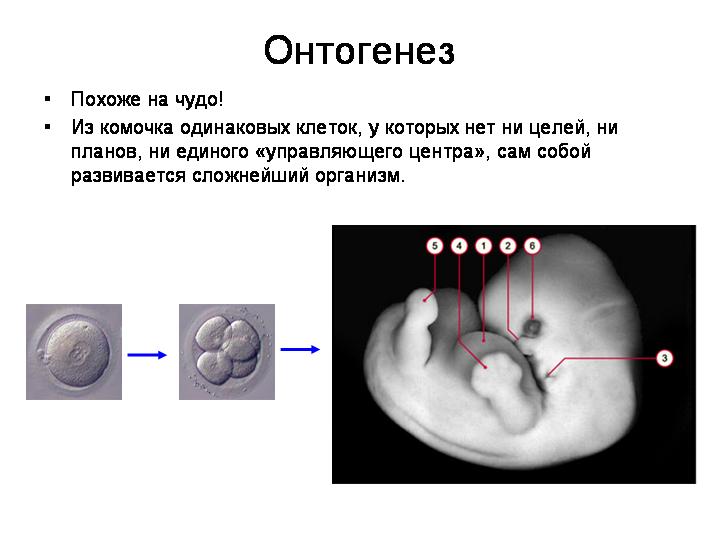
Принципиальный вопрос: как клетки зародыша «понимают», что одни должны стать печенью, другие костями, третьи мозгами? Казалось бы, должен быть какой-то «центральный управляющий орган», главный центр, руководящий развитием. Иначе – хаос. Где же этот центр?
Удивительно, но единого центра действительно нет. Оказывается, множество одинаковых объектов – например, делящихся клеток – может вести себя вполне «осмысленно» и формировать сложные структуры, не имея единого управляющего центра. Такие процессы называют самоорганизацией или самосборкой. Они всегда вызывают у нас ощущение чуда, чего-то нереального, но это – проблема нашей психики, а не объективной реальности.

Анекдот. Однажды Джон Холдейн, эволюционист, известный своими афоризмами, читал публичную лекцию, после которой одна дама встала и сказала: «А все-таки я не могу поверить, что из одноклеточного организма само собой могло развиться такое сложное и совершенное существо, как человек, даже если у него были на это миллиарды лет». Холдейн на это ответил: «Мадам, вы сами это проделали. И у вас ушло на это всего девять месяцев».
Мы могли бы на это возразить, что между онтогенезом и эволюцией (от одноклеточных к многоклеточным) есть принципиальная разница: в случае онтогенеза исходная клетка содержит некую генетическую «программу развития», сложившуюся в ходе предшествующей эволюции. В случае же эволюционного развития аналогичной программы нет.

Но если подумать, это различие хоть и существует, конечно, однако на самом деле оно, возможно, не настолько глубоко, как кажется на первый взгляд. Ведь на самом деле в зиготе нет «программы развития» многоклеточного организма, так сказать, в чистом виде. Если рассматривать геном как программу, то это на самом деле не программа развития эмбриона. Это программа поведения клетки, одной клетки, и не более того.
Все остальное – это самоорганизация, или самосборка.
Базовые принципы онтогенеза многоклеточных во многом контринтуитивны, трудны для понимания. Потому что мозг Homo sapiens плохо приспособлен для понимания процессов самоорганизации и самосборки. Наше мышление специализировано для целеполагания, мы привыкли планировать свои действия, ориентируясь на ожидаемый результат. Поэтому нам и кажется, что если чего-то не предусмотришь, не проконтролируешь, то ничего хорошего и не выйдет – только разруха и хаос.
Поэтому когда мы видим, как из чего-то простого само собой вдруг образуется что-то сложное, нам это кажется чудом, и мы начинаем подозревать воздействие какой-то особой организующей силы. Мы начинаем изобретать лишние сущности. Будь то Ламарковское «стремление к совершенству», «радиальная энергия» Тейяра де Шардена, «активность» Юрия Викторовича Чайковского, номогенез, ортогенез и так далее. Ну а в случае самосборки снежинки из хаотически движущихся молекул воды – совершенно очевидно вмешательство деда Мороза.
Где закодировано строение снежинки? Оно закодировано в свойствах молекулы воды в таком же смысле, в каком взрослый фенотип закодирован в геноме зиготы. И внешняя среда в обоих случаях сильно влияет на результат.
Контринтуитивность онтогенеза (как и эволюции) порождает множество проблем, и многие блестящие теоретики действительно спотыкались на этом месте.

Попробуем разобраться постепенно.
Вернемся к комочку клеток — зародышу, который образовался из яйцеклетки в результате нескольких первых делений. Каждая клетка зародыша имеет один и тот же геном. Геном определяет все свойства клетки, это ее «программа поведения». Программа у всех клеток зародыша одинаковая. Однако клетки начинают вести себя по-разному: одни превращаются в клетки кожи, другие — в клетки кишечника, и так далее.
Это происходит благодаря тому, что клетки обмениваются информацией — посылают друг другу химические сигналы и меняют свое поведение в зависимости от того, какие сигналы они получили от соседей. Кое-какие сигналы приходят и из внешнего мира. Например, клетки зародышей у растений чувствуют земное притяжение и принимают его в расчет, когда решают, как им себя вести. Наконец, яйцеклетка может с самого начала иметь простенькую «разметку»: один ее полюс может отличаться от другого по концентрации каких-нибудь веществ.

Программа поведения у всех клеток изначально одна и та же, но она может состоять из нескольких отдельных наборов правил. То, какой из наборов правил данная клетка будет выполнять, зависит от получаемых клеткой сигналов. Каждое отдельное «правило» выглядит примерно так: «если выполняются такие-то условия, сделай такое-то действие». Основные действия, которые делают клетки, —включение или выключение определенных генов. Это меняет свойства клетки, она начинает по-другому себя вести, по-другому реагировать на сигналы.
Правила поведения клетки «сделаны» из рецепторов, транскрипционных факторов, энхансеров, сигнальных молекул и белков, осуществляющих синтез этих молекул. Например, правило «если получен сигнал А, начни выделять вещество Б», может быть сделано из рецептора в-ва А, который активирует транскрипционный фактор В, который прикрепляется к энхансеру Г, расположенному около гена Д, который кодирует фермент, отвечающий за синтез вещества Б.
Как же получается, что клетки зародыша, имеющие одинаковую программу поведения и находящиеся, казалось бы, в одинаковых условиях, всё-таки ведут себя по-разному? Дело в том, что они на самом деле находятся в разных условиях — это получается само собой в процессе деления клеток. Кто-то оказался внутри, кто-то снаружи, кто-то снизу, кто-то сверху, в ком-то концентрация вещества А высокая (потому что данная клетка сформировалась из той части яйцеклетки, где этого вещества было много), а в ком-то вещества А мало.
Еще у клеток может быть «счетчик делений», который сообщает им, сколько раз яйцеклетка уже поделилась. Этот счетчик тоже химический: в яйцеклетке изначально были определенные вещества, запас которых не пополняется во время развития зародыша, и по тому, сколько в клетке осталось этих веществ, можно понять, сколько делений прошло с момента начала развития.
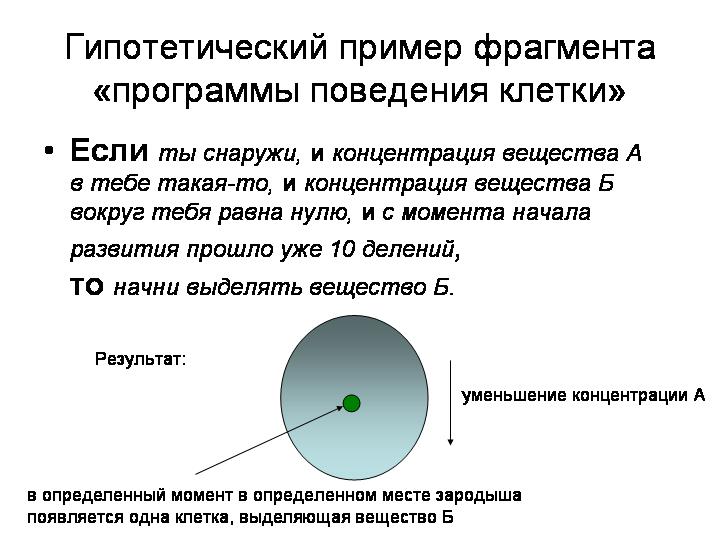
Программа поведения клетки может содержать, например, такие команды:
«Если ты снаружи, и если концентрация вещества А в тебе такая-то, и если концентрация вещества Б вокруг тебя равна нулю, и если с момента начала развития прошло уже 10 делений, тоначни выделять вещество Б».
К чему приведет выполнение такой команды? Оно приведет к тому, что на поверхности зародыша в определенный момент (после десяти делений) появится одна-единственная клетка, выделяющая вещество Б. Она будет расположена на строго определенном расстоянии от одного из полюсов зародыша, потому что в нашем примере вещество А служит для изначальной разметки яйцеклетки. По концентрации вещества А клетка может определить, на каком расстоянии от полюсов зародыша она находится.
Почему такая клетка, выделяющая вещество Б, будет только одна? А потому, что есть инструкция: «Если концентрация вещества Б вокруг тебя равна нулю». Как только первая клетка, в которой выполнятся поставленные условия, начнет выделять вещество Б, его концентрация перестанет быть равна нулю, и поэтому другие клетки не начнут его выделять.
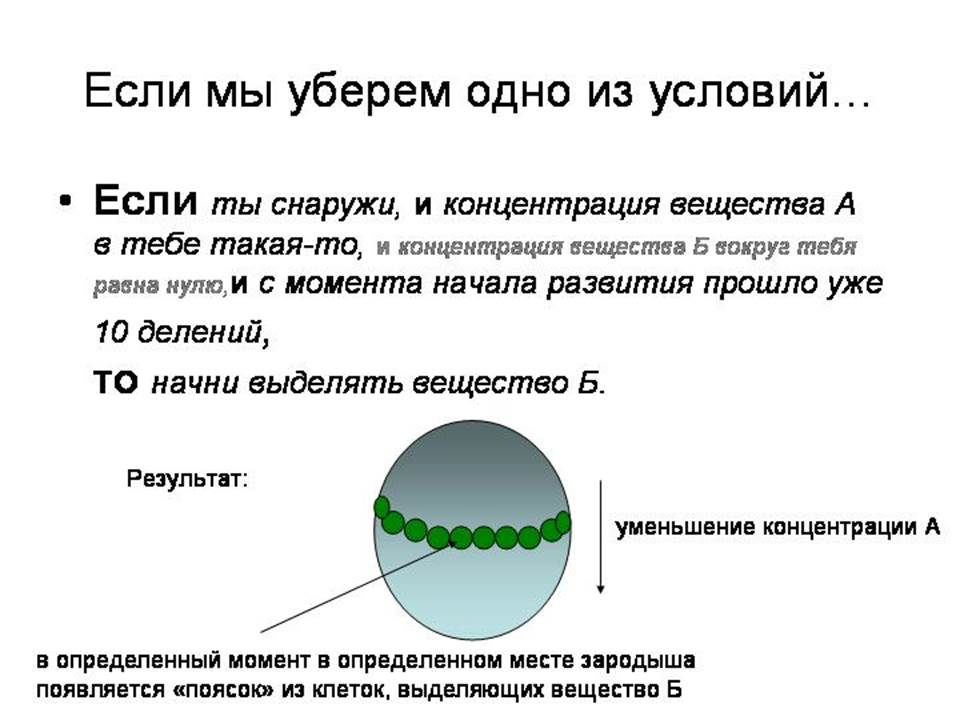
А что будет, если мы уберем из программы инструкцию «Если концентрация вещества Б равна нулю»? Тогда вещество Б начнет выделяться не одной клеткой, а целой полоской клеток, опоясывающей зародыш на определенном расстоянии от полюсов. Ширина пояска и его положение (ближе или дальше от того полюса, где концентрация А максимальна) будут зависеть от того, какие концентрации вещества А указаны в инструкции «Если концентрация вещества А в тебе такая-то».
Теперь наш зародыш размечен уже гораздо сложнее и интереснее, чем раньше. У него есть «передняя часть», в которой много А, а концентрация Б растет спереди назад; у него есть центральный поясок, где максимальна концентрация Б; и у него есть задняя часть, где мало А и где концентрация Б убывает спереди назад. Наш зародыш подразделился на четко отграниченные друг от друга части, в которых клетки находятся в разных условиях и поэтому будут выполнять разные подпрограммы своей исходной общей программы.
Мы подразделили зародыш на передний, средний и задний отделы. Они могут стать, например, головой, туловищем и хвостом. Но хотелось бы еще понять, где у него будет спина, а где живот. Как это сделать? Очень просто, мы уже это проходили. Нужна инструкция, приводящая к появлению одной клетки или группы клеток, выделяющих какое-нибудь вещество (например, В) на любом «боку» зародыша, где-нибудь посередине между головой и хвостом. И пусть это вещество В запустит программу развития спины там, где его много, и программу формирования брюха там, где его мало. Когда зародыш подробно «размечен», каждая группа клеток может без труда определить, где она находится, и активизировать заготовленную на этот случай подпрограмму (набор правил поведения).
В ходе развития зародыша действительно то тут, то там появляются особые «управляющие центры» или организаторы — группы клеток, выделяющие то или иное вещество, которое служит для других клеток сигналом и влияет на их поведение. Но при этом все клетки по-прежнему ведут себя в строгом соответствии с изначальной генетической программой, которая у всех одна и та же. Управляющие центры возникают сами, путем самоорганизации, никто их нарочно туда не вставляет. И никакого «единого централизованного руководства» для этого не требуется и его нет.
В развитии настоящих животных всё сложнее, чем в нашем воображаемом примере, но, как ни странно, ненамного.

Итак, основной принцип, лежащий в основе онтогенеза многоклеточных, состоит в том, что онтогенез – это процесс самосборки упорядоченных многоклеточных структур, формирующихся за счет согласованного поведения множества индивидуальных модулей (клеток), причем все эти модули изначально следуют одному и тому же набору «правил поведения», закодированному в геноме.
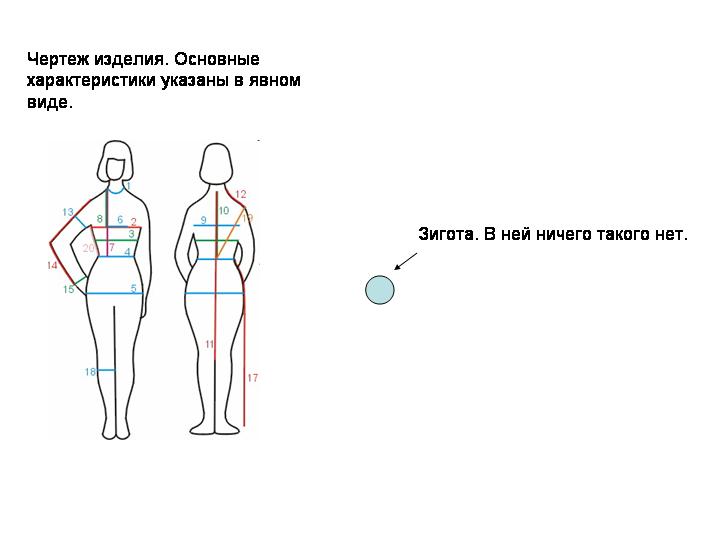
Еще несколько пояснений. Ведь мы говорим о вещах действительно контринтуитивных. Геном зиготы не содержит ничего похожего на «чертеж» организма. Между элементами чертежа и элементами изделия, как правило, существует соответствие «один к одному». Каждый элемент чертежа соответствует определенному элементу изделия и наоборот. В геноме зиготы ничего такого нет. Между частями (признаками) организма и частями генома (генами, локусами) нет соотношения «один к одному»: большинство признаков зависит от множества генов и наоборот.
Чуть точнее будет сравнение с рецептом (или алгоритмом), на основе которого повар готовит торт. Между частями пирога или соуса и отдельными элементами рецепта (словами, предложениями) нет взаимно однозначного соответствия. По готовому изделию можно восстановить его чертеж близко к оригиналу. Восстановить по «фенотипу» пирога его рецепт принципиально труднее, а восстановить его дословно в общем случае невозможно. Но метафора «рецепта» тоже несовершенна. Рецепт содержит команды на уровне целого изделия («придать тесту такую-то форму», «тщательно перемешать»). Но в геноме зиготы нет инструкций, выполняемых на уровне целого эмбриона. Геном содержит лишь набор локальных правил поведения, выполняемых на уровне отдельных клеток. Геном можно называть рецептом или алгоритмом только с этой оговоркой.
В программировании такие алгоритмы хорошо известны. Их называют «алгоритмами самосборки».
Если бы развитие шло не путем самоорганизации на основе программы, а по чертежу, нам было бы труднее эволюционировать. Лет сто назад, когда мы еще не знали законов развития эмбриона, многое в эволюции казалось непонятным. Например, некоторые удивлялись, как могут в процессе эволюции удлиниться все четыре ноги одновременно — ведь для этого нужно, чтобы мутации одновременно изменили длину сразу всех четырех ног! Действительно, если бы в геноме был записан чертеж организма, то потребовалось бы внести в этот чертеж целых четыре поправки, чтобы увеличить длину четырех ног. Теперь-то мы знаем, что развитие идет по программе, в которую достаточно внести всего одно изменение, чтобы длина всех четырех конечностей изменилась, причем изменилась одинаково.
Математики говорят, что закодировать в геноме чертеж животного было бы намного сложнее, чем такую программу. Эта программа, как ни странно, сама по себе гораздо проще, чем получающийся в результате организм.
Итак, геном кодирует интерактивную программу поведения клетки. В основе этой программы лежат генно-регуляторные сети, расшифровкой которых занимается эволюционная биологии развития, так называемая эво-дево.
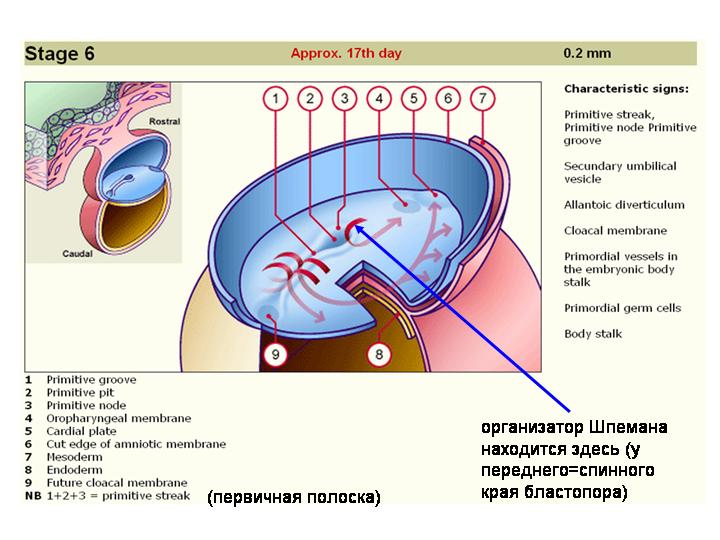
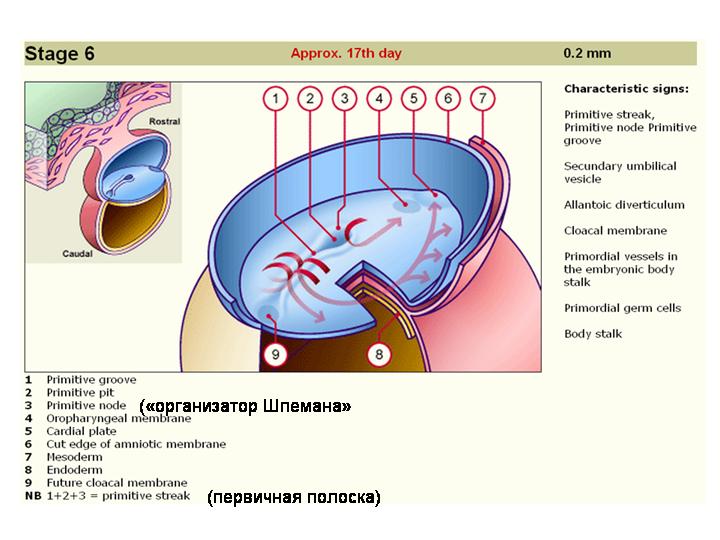
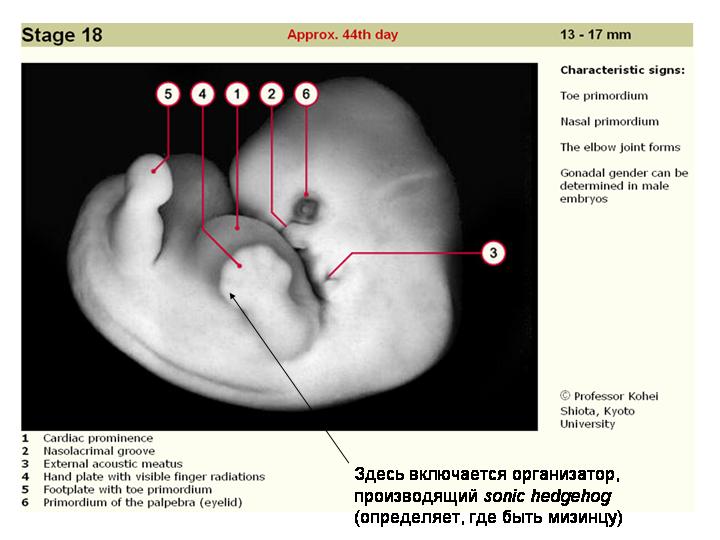
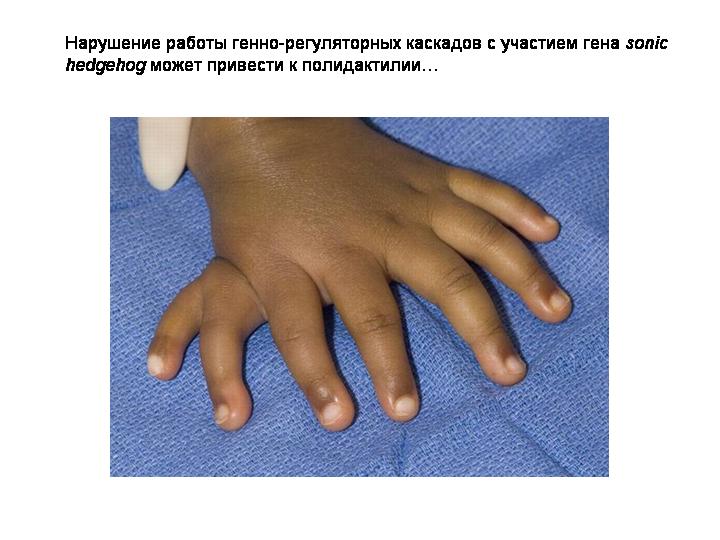
Вот где расположен организатор Шпемана у человеческого эмбриона.
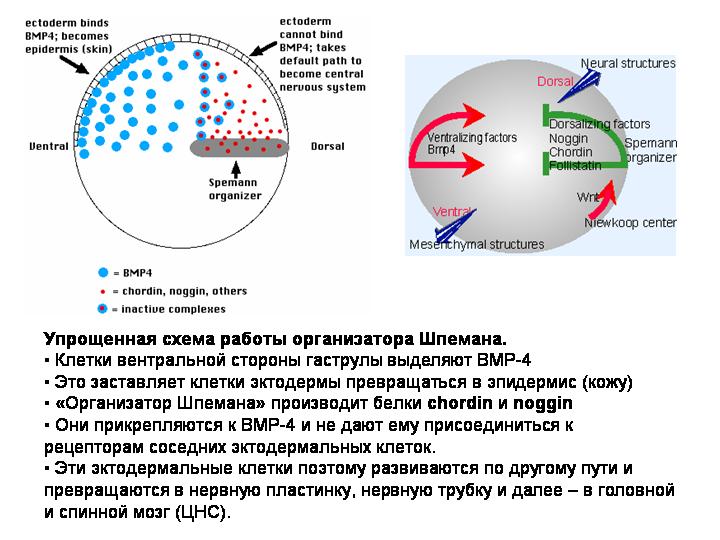
ГЕННО-РЕГУЛЯТОРНЫЕ КАСКАДЫ
Ход онтогенеза определяется генно-регуляторными сетями (каскадами). В них участвуют сигнальные белки и др. в-ва («морфогены»; выделяются клеткой в окружающее межклеточное пространство), рецепторы, транскрипционные факторы, малые регуляторные РНК. Энхансеры (сайты связывания ТФ) в регуляторных областях генов-регуляторов – важный компонент «генетической программы развития». От энхасеров зависит, какими переключателями (а значит, где и когда) будет включаться данный ген.
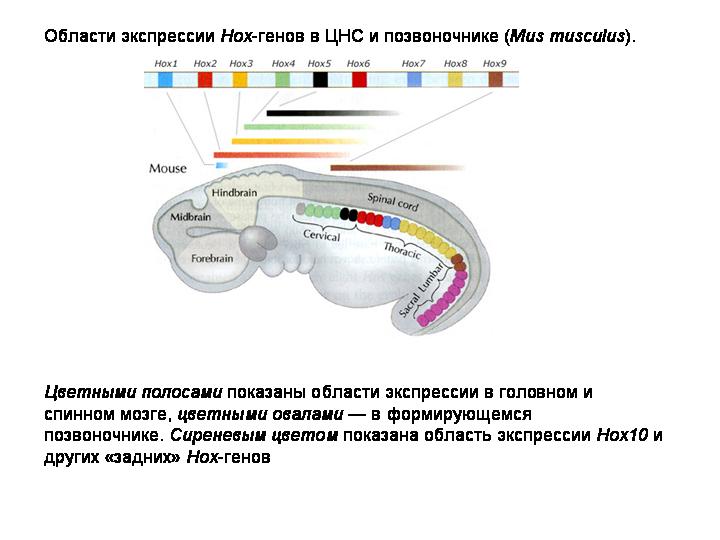
ХОКС-ГЕНЫ
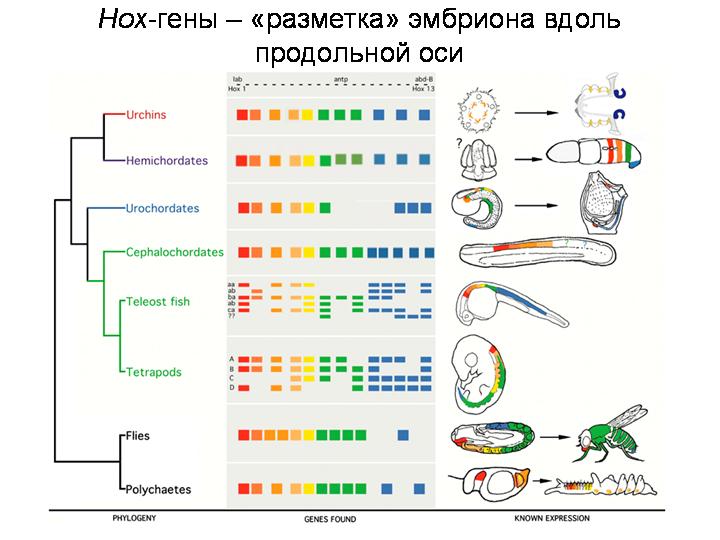
У всех животных за разметку эмбриона вдоль передне-задней оси отвечает особое семейство генов – HOX-гены. Сначала нашли у дрозофилы, затем у всех животных.
Открытие сходных Hox-генов у разных типов животных заставило по-новому взглянуть на морфогенез животных и его преобразования в ходе эволюции. Стало ясно, что, изменив один ген или время (или место) его включения, можно трансформировать, создать, удалить или перенести в другое место сразу целый орган, сохранив при этом общий план строения.
Hox-гены у дрозофилы, человека и многих других животных располагаются в хромосоме в строгом порядке, в том самом, в котором происходит дифференцировка основных частей тела двусторонне симметричного животного. Сначала у раннего эмбриона начинают работать гены, отвечающие за строение органов на голове, затем на груди, затем гены начинают оформлять и хвостовую часть.

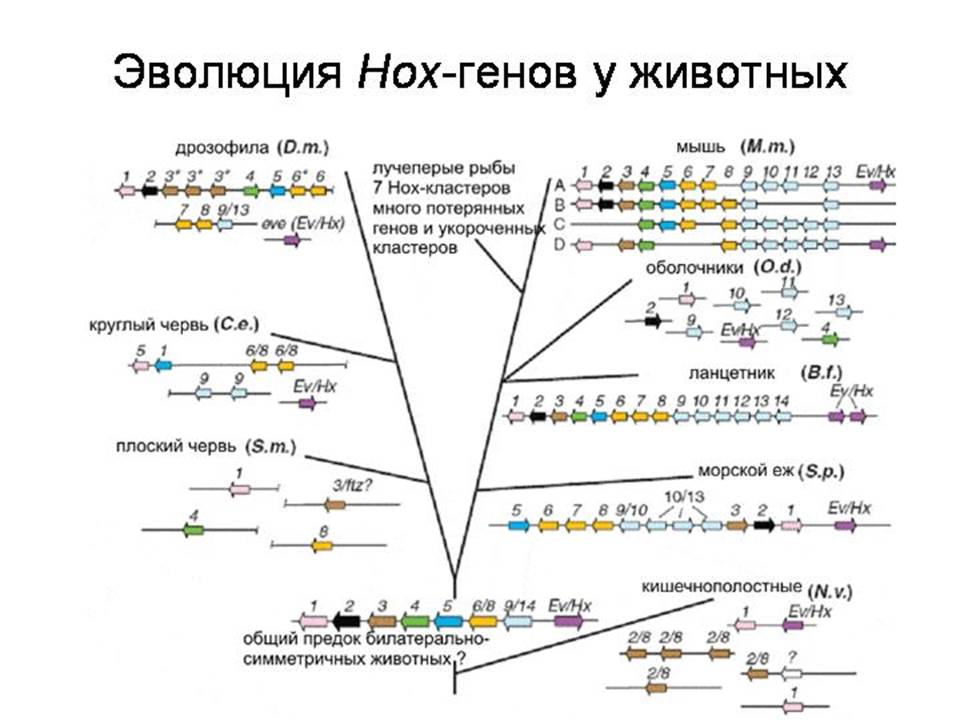
Семейство Hox-генов подразделяется на 14 классов. Считается, что эти 14 классов возникли путем дупликации одного или немногих исходных генов, реплики затем мутировали и обретали новые функции. У кишечнополостных и гребневиков имеется всего 4 класса Hox-генов.
У предполагаемого общего предка двустороннесимметричных животных их должно было быть по крайней мере 8. У млекопитающих присутствуют все 14 классов.
Принцип работы этих генов одинаков. Все они являются транскрипционными факторами, то есть их функция состоит во «включении» или «выключении» других генов. В результате работы Hox-генов запускается каскад реакций, приводящий к появлению в клетке нужных белков.
Позже выяснилось, что у некоторых животных они расположены вовсе не так правильно, как у человека и дрозофилы.

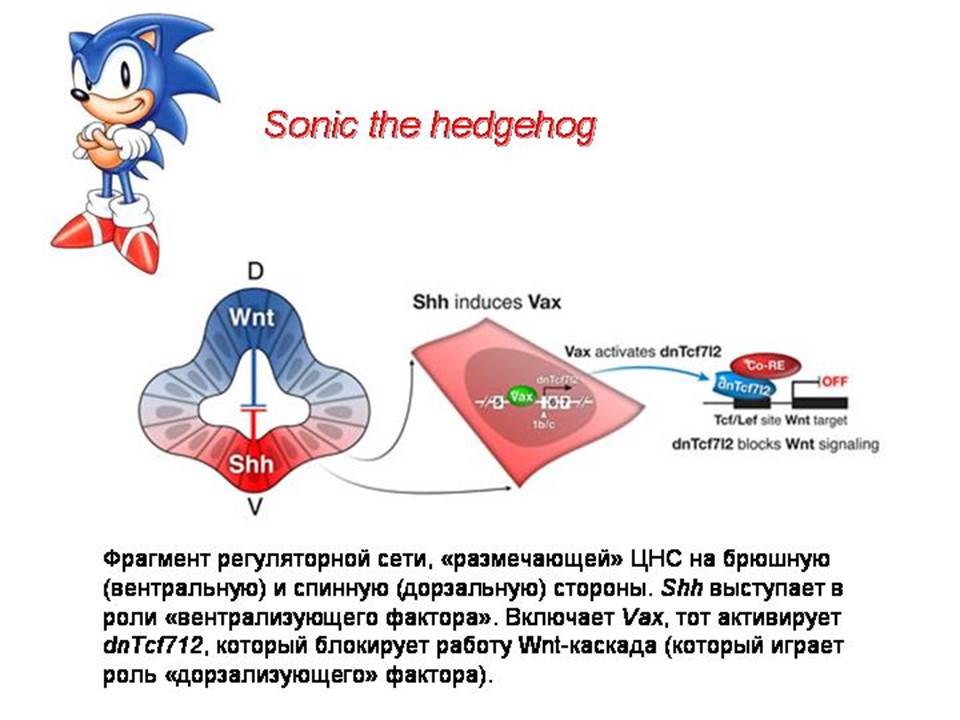
Кроме Хокс-генов, существует много других регуляторов развития. Для большинства характерна плейотропность. Плейотропность – множественность функций и фенотипических проявлений. Один и тот же ген-регулятор (ТФ) может регулировать несколько совершенно разных процессов на разных стадиях эмбрионального развития. Это «профессиональные переключатели», которым в принципе все равно, что переключать (был бы у регулируемого гена нужный энхансер). Поэтому в ходе эволюции под их управление легко могут попасть новые «подпрограммы». Так возникают новые признаки.
Модель

Итак, онтогенез - это процесс самоорганизации, в ходе которого из согласованных действий множества одинаково запрограммированных клеток, следующих сравнительно простому набору правил поведения, «самозарождаются» сложные многоклеточные структуры.
Какими свойствами должен обладать онтогенез многоклеточных исходя из того, что он основан на этом принципе? Похоже на то, что многие специфические, нетривиальные свойства онтогенеза могут быть на самом деле неизбежными следствиями этого принципа. В таком случае для них не нужны специальные объяснения.
Мы попытались при помощи моделирования определить круг свойств онтогенеза, которые могут быть следствиями этого принципа. Для этого была разработана программа EvoDevo.
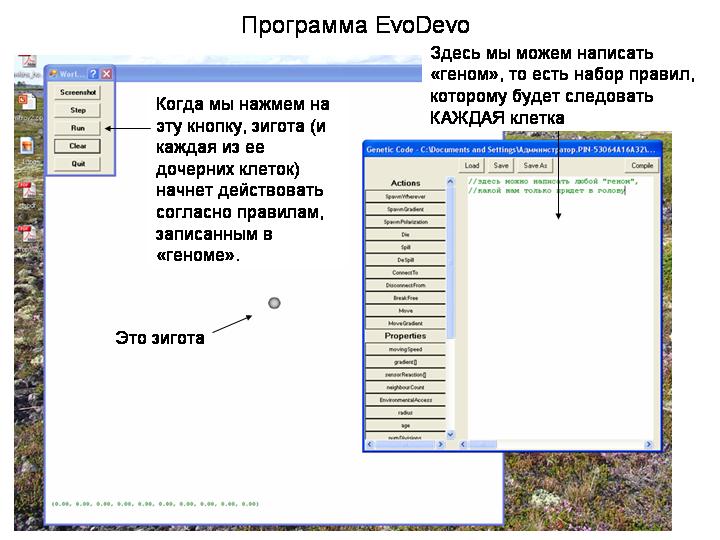
Есть такой афоризм – кстати, совершенно неправильный – что модель, это такая штука, в которую что заложишь, то и получишь. Так вот, в программу Эво-Дево изначально НИЧЕГО не заложено, кроме вот этого принципа.
Программа дает нам одну исходную клетку – зиготу. Мы можем задать любой набор правил поведения клетки. Следовать этим правилам будут все клетки нашего эмбриона. Мы не можем задать правило для всего эмбриона. Мы не можем сказать бластуле «сделай себе впячивание и превратись в гаструлу». Мы можем задать только правила для КЛЕТКИ, например: при таких-то условиях мигрируй туда, где возрастает концентрация такого-то вещества. Причем набор правил один и тот же для всех клеток. Мы пишем этот набор правил вот в этом окошке, потом нажимаем кнопку RUN, и наша зигота начинает выполнять наши правила.
Клетка может выполнять следующие действия: выделять одно или несколько сигнальных веществ («морфогенов»), Делиться, Перемещаться, Отмирать, Становиться клеткой того или иного «типа» (аналог клеточной дифференциации) и т.д.
Каждое действие выполняется (или не выполняется) клеткой в зависимости от комбинации указанных в «генотипе» условий, таких как концентрация морфогенов, число клеток-соседей и т.д.
Программа позволяет, произвольно меняя «генотип», создавать неограниченное разнообразие онтогенезов и «организмов».
Вот несколько примеров тварюшек.
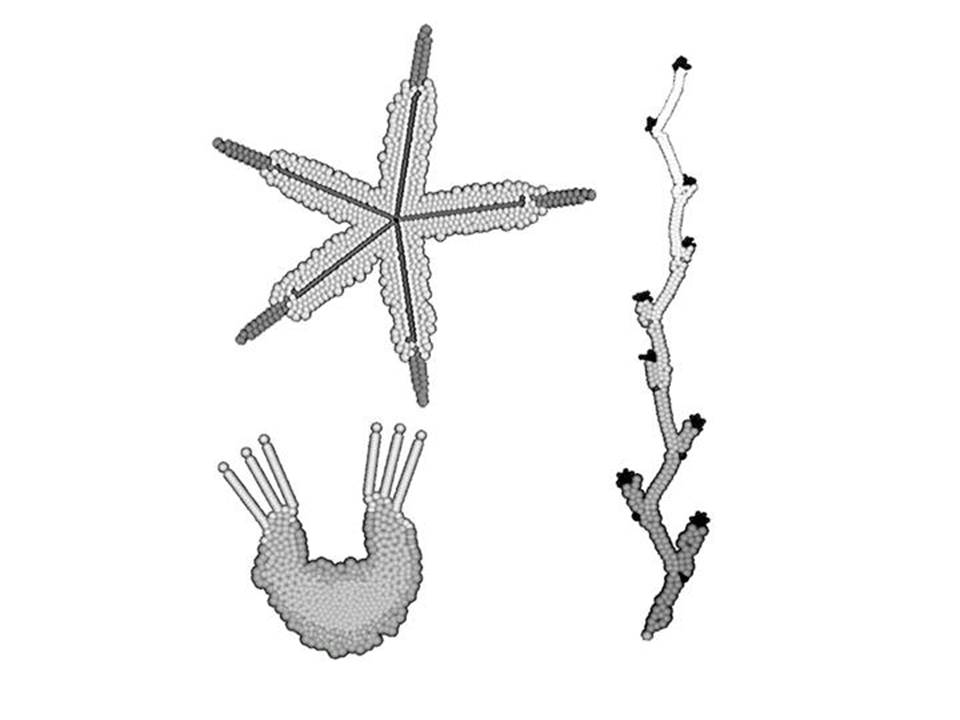

Этот генотип мы рассмотрим в качестве примера поподробнее. Генотипы у нас пишутся, конечно, не в виде последовательностей нуклеотидов, а на более высоком уровне, который примерно соответствует уровню генно-регуляторных сетей. Это уровень поведения клетки, включая ее реакции на то, что происходит вокруг.
Вот пример генотипа. Вы видите, что это набор неких правил поведения клетки. В действительности генотипы у нас пишутся на языке С++. Но здесь для удобства я перевел этот генотип на русский язык.
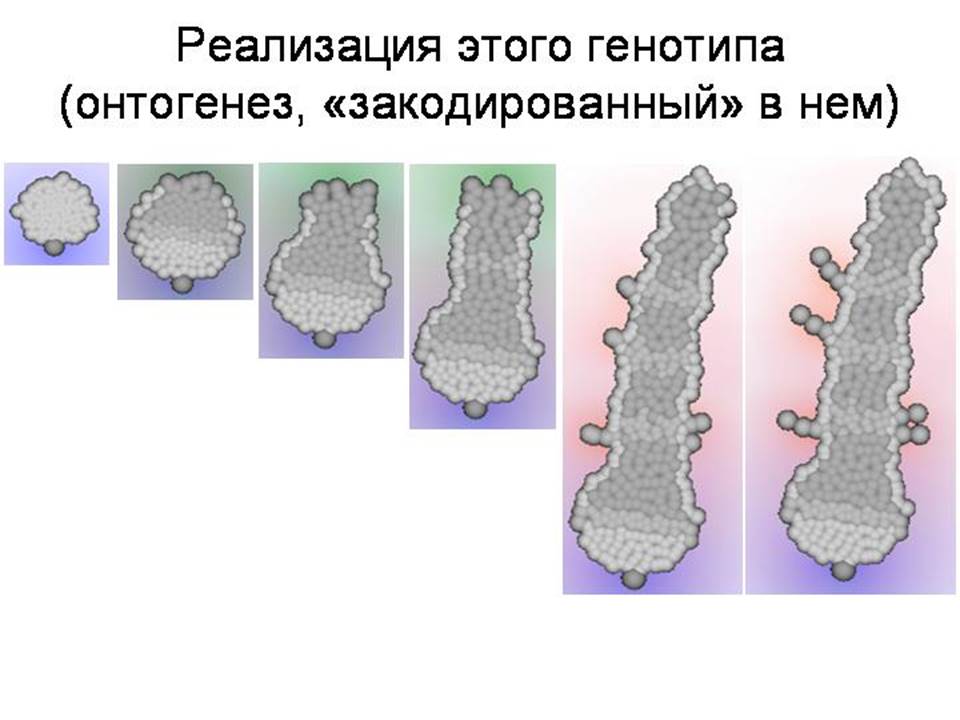
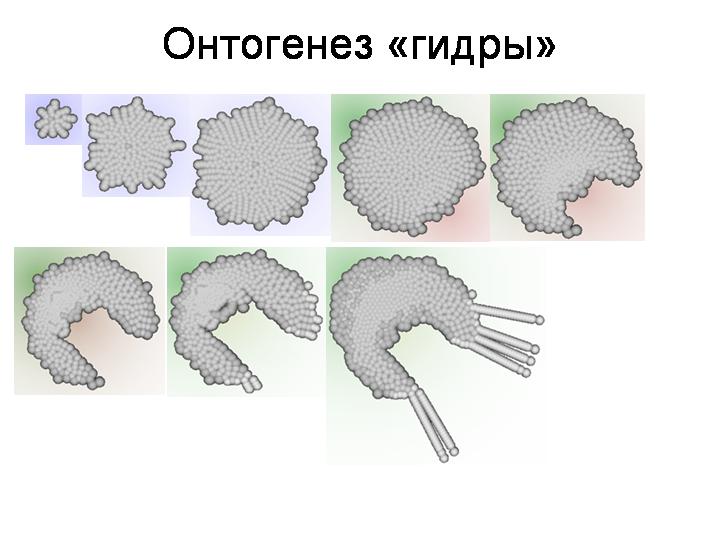
Онтогенез, генерируемый этим генотипом, показан на этом слайде. Можно заметить, что для понимания соотношения между генотипом и фенотипом у модельных организмов, как и у реальных животных, требуются известные усилия. Глядя на генотип, трудно предсказать, какой фенотип будет им сгенерирован. Тем не менее, в отличие от реальных животных, в модели все это можно проследить и понять.
ДЕМОНСТРАЦИЯ ПРОГРАММЫ. Пояснение, как развивается червяк. В данном случае онтогенез протекает следующим образом. Сначала в результате неупорядоченных делений исходной клетки и ее потомков формируется округлое скопление клеток. Затем одна из клеток на краю скопления начинает выделять морфоген 0, который задает передне-заднюю полярность эмбриона. Эта клетка становится «организатором», маркирующим передний конец эмбриона. Когда скопление немного увеличится в размере за счет роста клеток, на его противоположном конце возникает второй «организатор» – клетка, выделяющая морфоген 1 и маркирующая задний конец эмбриона. После этого происходит дифференцировка клеток эмбриона на клетки передней и задней полусфер и «пояска»; этот процесс управляется соотношением концентраций морфогенов 0 и 1. Внешние клетки передней полусферы образуют плотный «эпителий». Клетки пояска начинают выделять морфоген 2, который ингибирует деление клеток задней полусферы. Внешние клетки задней полусферы, достаточно удаленные от заднего организатора («боковые»), превращаются в клетки типа 1 и образуют плотный эпителий, фиксируя форму растущего червя. Внешние клетки задней полусферы, достаточно удаленные от пояска и достаточно близкие к заднему организатору («хвостовые»), делятся, обеспечивая нарастание заднего конца эмбриона. Внутренние клетки задней полусферы, достаточно удаленные от заднего организатора и от источников морфогена 2, превращаются в клетки септ, разделяющих «сегменты», и начинают выделять морфоген 2 (то же самый, что и клетки пояска). Когда тело червя достигает определенной длины, концентрация морфогена 0 становится достаточно низкой, и тогда задний организатор прекращает выделение морфогена 1, что ведет к прекращению роста эмбриона и образованию плотного эпителия на его заднем конце. После этого все клетки септ однократно делятся, в результате чего часть этих клеток мигрирует («продавливается») сквозь эпителий и вступает в контакт к внешней средой, что приводит к их превращению в клетки типа 5. Последние начинают выделять морфоген 3 и делятся по градиенту морфогена 2, что ведет к росту «конечностей», который лимитируется концентрациями морфогенов 2 и 3.
У модельных организмов обнаруживается ряд важных свойств, похожих на то, что наблюдается в реальных онтогенезах.

Первое свойство – это стохастичность. Если не ввести в геном специальных стабилизирующих инструкций, то, какого бы зверя мы ни создали, фенотип будет неустойчив. Например, данный генотип не всегда дает «нормального» сегментированного червя: развитие часто останавливается на ранней стадии.
По-видимому, онтогенезу реальных организмов тоже присуща такая стохастичность, которая, однако, обычно не проявляется из-за наличия специальных стабилизирующих адаптаций (помните, мы говорили о помехойстойчивости?).
Всегда есть неизбежные флуктуации на уровне биохимии и экспрессии генов. Активность гена невозможно отрегулировать с абсолютной точностью. Поэтому две клетки с одинаковыми геномами обязательно будут различаться по числу молекул тех или иных белков. Это ведет к различиям в поведении клеток.
Как правило, в модели не удается уменьшить стохастичность онтогенеза, не усложняя геном, т.е. не внося в него дополнительных правил.
Введение дополнительного правила при моделировании аналогично появлению в ходе эволюции нового энхансера, ставящего активность гена – регулятора развития в зависимость от какого-то дополнительного фактора. По мнению ведущих специалистов в области Evo-devo, именно изменения регуляторных участков ключевых генов – регуляторов развития играют основную роль в морфологической эволюции животных.
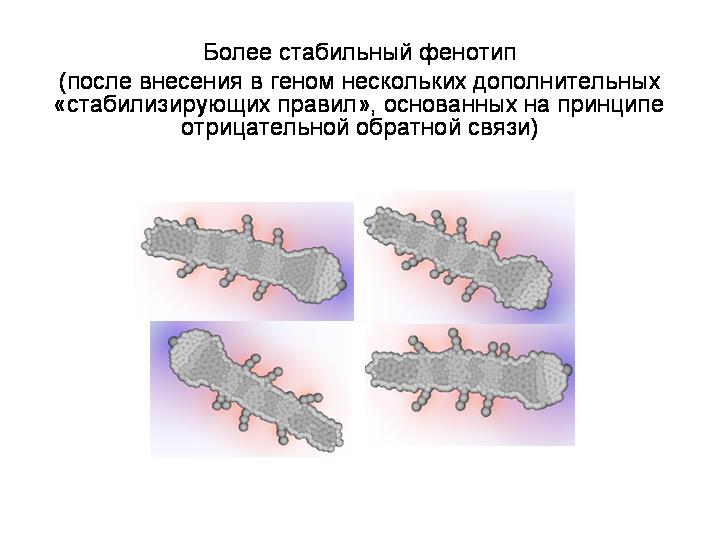
Мы стабилизировали этого червяка, добавив несколько правил. С этим «стабилизированным» генотипом мы провели «генно-инженерные эксперименты». Случайные мутации вносили в генотип по одной и смотрели, как эта мутация отразится на фенотипе. Были выявлены следующие закономерности:

Наличие в программе развития отрицательных обратных связей является необходимым условием для формирования сколько-нибудь сложного, упорядоченного фенотипа. В онтогенезе нашего червячка на обратных связях построена, например, система выделения клетками морфогенов. Клетки начинают выделять морфоген только если его концентрация достаточно низка, и не выделяют его, если он уже выделяется в достаточном количестве другими клетками.
Утрата таких обратных связей приводит к краху системы онтогенеза, т.е. к резкому снижению способности к самоорганизации. Фенотип получается радикально упрощенным или разупорядоченным, дезорганизованным.
Для стабилизации признака, нестабильно воспроизводящегося в онтогенезе, часто нельзя обойтись без усложнения генетической программы развития, без добавления в нее дополнительных регуляторных контуров.
Это согласуется с идеями о «творческой роли» стабилизирующего отбора (Шмальгаузен). Отбор на стабильность должен вести к усложнению программы развития. То есть отбор, просто отсеивающий «уродцев», в перспективе способствует развитию новых генно-регуляторных контуров, повышающих стабильность воспроизведения «нормального» фенотипа.
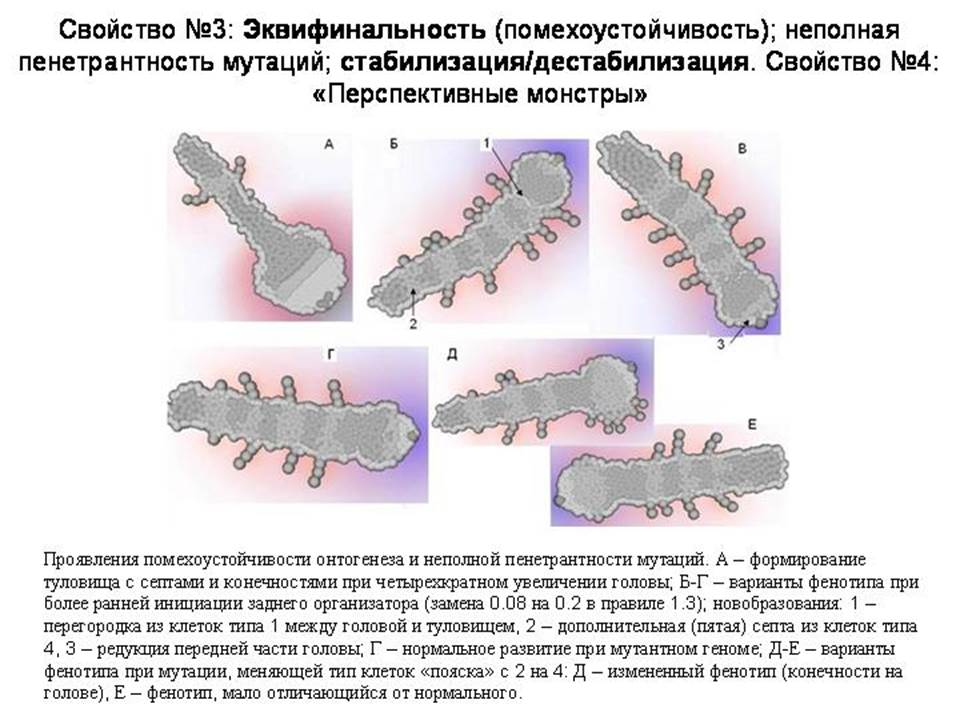
Эти неизбежно возникающие отрицательные обратные связи и специальные стабилизирующие механизмы придают онтогенезу помехоустойчивость (или эквифинальность – т.е. одинаковый конечный результат при разных нарушениях в ходе развития). ООС позволяют онтогенезу компенсировать разные непредвиденные помехи и приходить, невзирая на них, к более или менее «нормальному» итоговому фенотипу. Фокус тут, по-видимому, в том, что регуляторный контур, развившийся для компенсации каких-то помех, возникающих по некоторой одной причине, будет с тем же успехом компенсировать такие же помехи, возникающие по любым другим причинам. В том числе – по причинам, которые не были изначально предусмотрены программистом (ну или естественным отбором).
На этом слайде показаны некоторые примеры. А – резко изменены ранние стадии развития, но тело с сегментами и придатками все равно сформировалось. Здесь было увеличено число делений на ранних стадиях развития, что непосредственно приводит к увеличению головы. Полученный результат можно также рассматривать как проявление 1) «плейотропности» (множественности эффектов) генов – регуляторов развития: одна мутация приводит одновременно и к увеличению головы, и сокращению числа сегментов туловища; 2) «морфогенетических корреляций»: увеличение головы автоматически приводит к олигомеризации туловища. То есть создается впечатление, что развивающийся модельный организм ведет себя как нечто целое (труды И.И.Шмальгаузена).
Многие мутации оказывают комплексное влияние на фенотип. По-видимому, плейотропность неизбежно вытекает из того обстоятельства, что клетки эмбриона влияют друг на друга и общаются при помощи химических сигналов. Поэтому поведение клеток оказывается взаимосвязанным, а весь организм ведет себя в определенном смысле как единое целое. Плейотропность генов – регуляторов онтогенеза – это один из важнейших эмпирических выводов современной эволюционной биологии развития (Carroll, 2008). Плейотропность связана и с тем обстоятельством, что имеющиеся морфогены являются очень удобными «заготовками» для формирования новых элементов генно-регуляторных сетей. Для того, чтобы создать новую упорядоченную структуру (например, дополнительную пару конечностей или пятно на крыле), как в реальной эволюции, так и в модели, проще не создавать «с нуля» новые генно-регуляторные контуры, а «пристроить» программу формирования новой структуры к уже имеющимся контурам. Отсюда неизбежно следует плейотропность.
Б, В Г – это фенотипические результаты одной и той же мутации. В первых двух случаях она привела к появлению интересных новшеств. Появилась септа нового типа между головой и туловищем, появился один дополнительный сегмент, и редуцировался один из секторов головы. То есть некоторые мутации приводят к появлению морфологических новшеств, причем степень сложности и упорядоченности фенотипа при этом не снижается или даже увеличивается.
Но в третьем случае при таком же геноме развился вполне нормальный фенотип. Это пример помехоустойчивости или, что то же самое, неполной пенетрантности, неполной проявляемости мутаций. Мутация может проявиться в фенотипе, а может и не проявиться. По-видимому, чем больше отрицательных обратных связей и стабилизирующих механизмов, тем выше вероятность, что случайная мутация не проявится в фенотипе или проявится слабо.
Д-Е: это другая мутация. В одном случае появился новый признак – придатки на голове, в другом – получился нормальный фенотип. Это два варианта развития при одном и том же геноме. Опять мы видим неполную пенетрантность нового признака, возникшего в результате мутации.
Эти же примеры показывают, что мутации могут приводить к дестабилизации онтогенеза. Исходный вариант генома никогда не приводил к придаткам на голове. Мутация может дестабилизировать онтогенез таким образом, что иногда все будет нормально, а иногда будут вырастать придатки на голове. Появление мутации с неполной пенетрантностью и дестабилизация онтгогенеза – это одно и то же. Был один стандартный фенотип – произошла мутация – получилось несколько вариантов фенотипа, один из которых соответствует старой норме, а другие представляют собой разные отклонения (морфозы). В результате мутации новый признак может возникнуть в виде редкой аномалии. Последующие мутации могут этот новый признак стабилизировать.
Неполная пенетрантность новых признаков, в сочетании с сохранением прежней «нормы» в качестве одного из вариантов развития, повышает эволюционную пластичность онтогенеза. Это свойство делает мутации более «безопасными». Редкая аномалия может оказаться вредной в одних условиях, но полезной в других, и тогда отбор поддержит мутации, повышающие частоту реализации данной аномалии, и она в итоге станет «нормой».
По-видимому, наличие в «генетической программе развития» обратных связей и стабилизирующих механизмов придает ей свойство, которое компьютерщики неформально называют «умом»: «умная» программа способна справиться с разными нештатными ситуациями, в том числе даже такими, которые не были предусмотрены программистом.
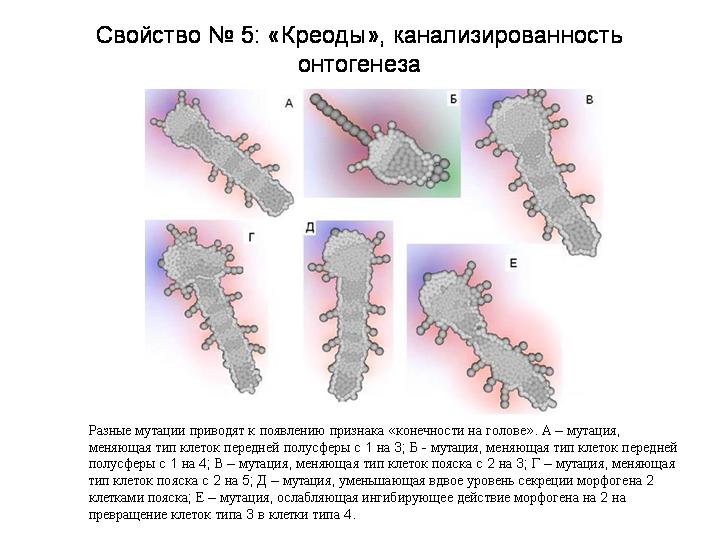
Одно и то же морфологическое изменение может быть вызвано многими разными мутациями. Один из примеров у нашего червяка – это появление ног на голове. Мы нашли 7 разных мутаций, приводящих к появлению этого признака. В результате морфологическое разнообразие, порождаемое мутациями, оказывается в целом гораздо меньше, чем разнообразие самих мутаций. Иными словами, в изменениях фенотипа оказывается намного меньше степеней свободы, чем в изменениях генотипа. Это позволяет говорить о «канализированности» эволюционных преобразований онтогенеза, о наличии в нем ограниченного числа скрытых альтернативных путей, или креодов. То есть в онтогенезе наших модельных зверей мы видим то же, что Уоддингтон и Шмальгаузен увидели в онтогенезе реальных животных.

Итак, многие свойства онтогенеза модельных организмов характерны и для онтогенеза реальных животных. На слайде перечислены такие свойства. Это позволяет предполагать, что заложенный в программу базовый принцип может быть достаточным объяснением этих свойств.
Этот принцип, напомню, состоит в том, что в ходе онтогенеза самосборка упорядоченных структур происходит благодаря согласованному поведению индивидуальных клеток, изначально «запрограммированных» одинаковым образом, причем в поведении этих клеток присутствует неустранимый элемент случайности.
ДОПОЛНЕНИЕ: Демонстрация некоторых других мутантов
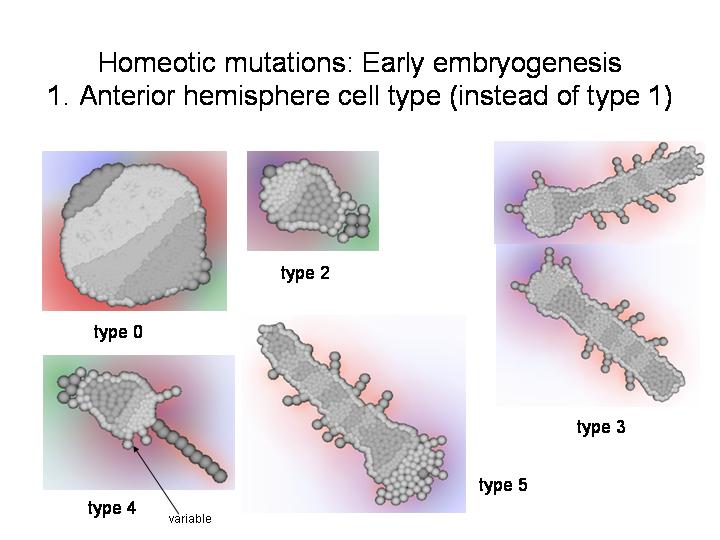
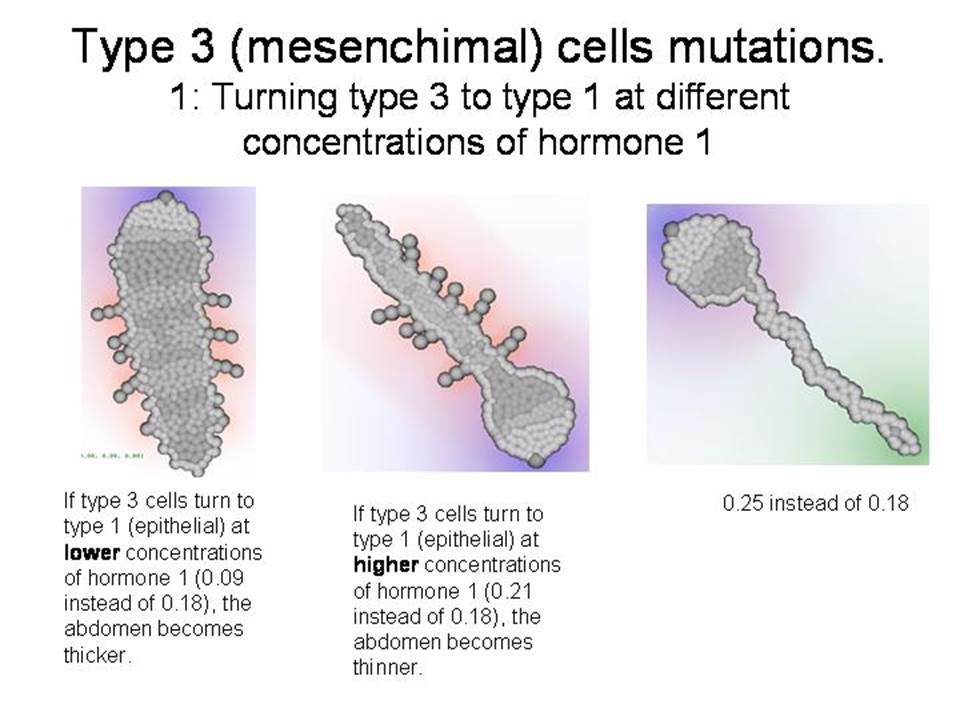
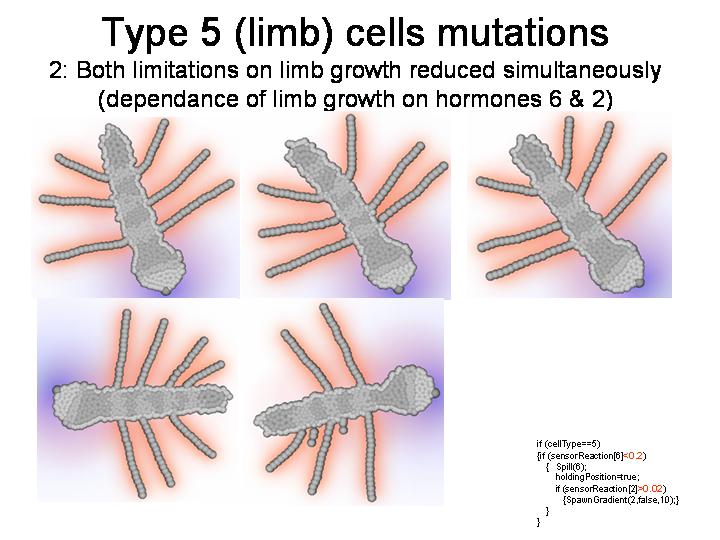



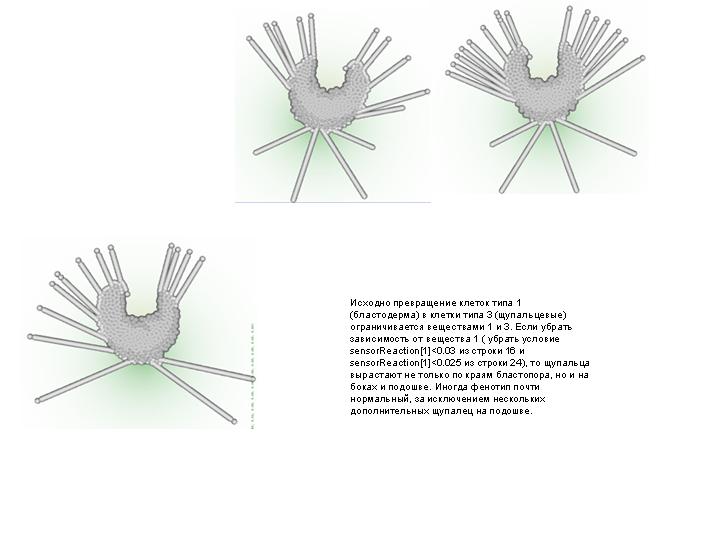
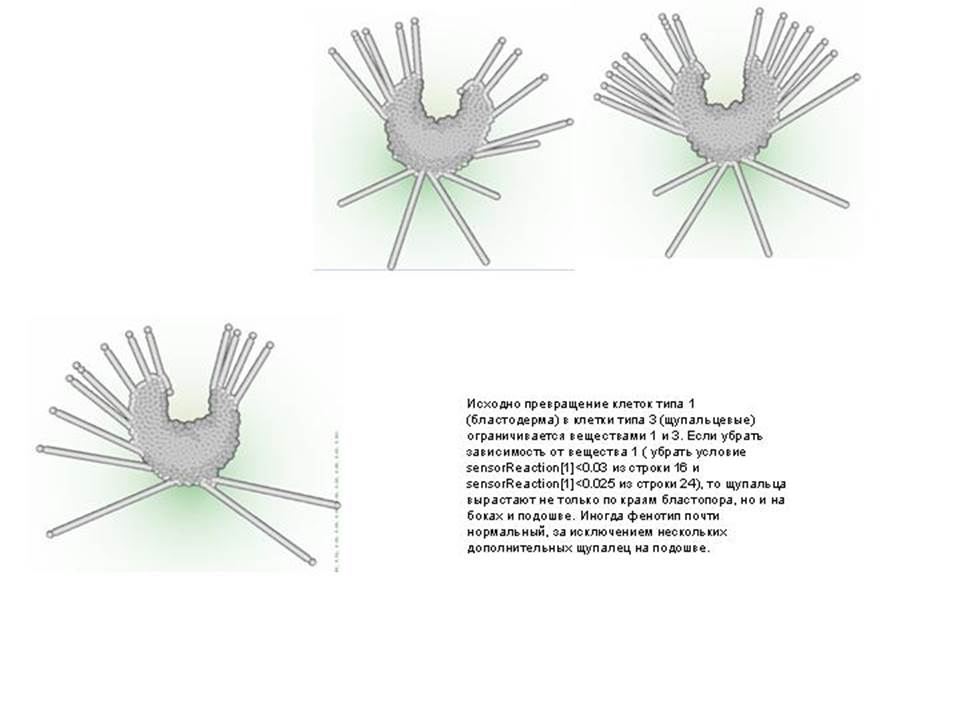
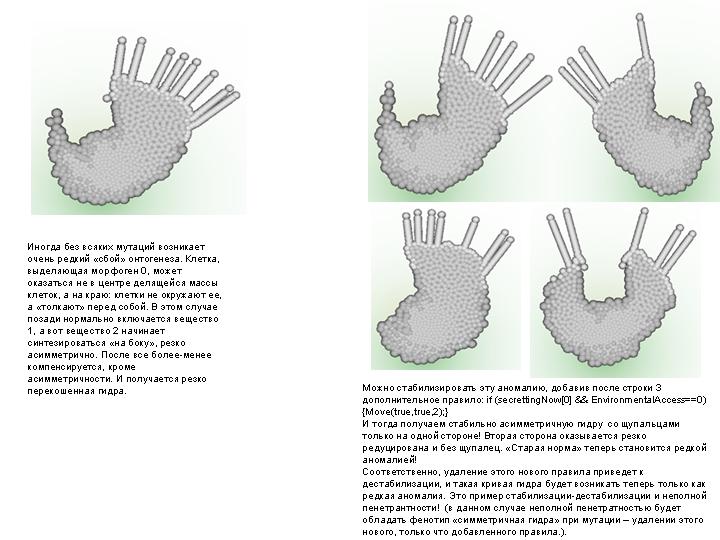
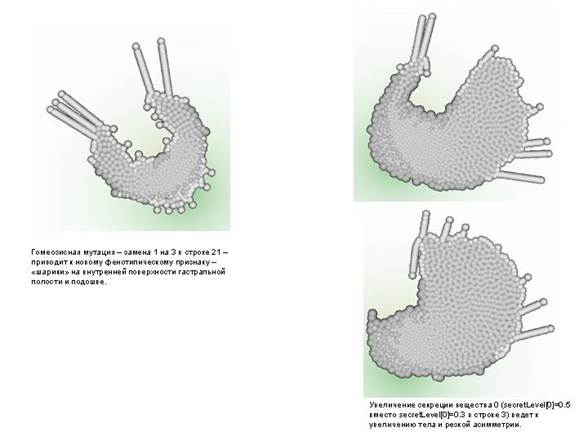
--------------------------------------------------------
16-е занятие: Письменная контрольная работа. 17-е занятие: разбор эссе и контрольной.
Часть 6. Происхождение жизни (лекции 18-20)



ВЕРОЯТНОСТИ. Одна авиакатастрофа примерно на 106 часов полета. Даже если мы летаем очень много, допустим, по 102 часов в год, то за свою жизнь налетаем не более 5000 часов. Вероятность погибнуть в авиакатастрофе всего лишь 1/200 (пренебрежимо мало). Но если бы мы жили по 100000 лет, то за жизнь летали бы 107 часов, и практически гарантированно погибали бы в авиакатастрофе.

Путь от неорганических молекул к первой живой клетке был долгим и трудным. За один шаг такие превращения не происходят. На этом долгом пути было множество развилок, остановок, временных отступлений, всевозможных промежуточных этапов и «переходных звеньев». Досаднее всего, что эти события не оставили, по-видимому, никаких следов в палеонтологической летописи. Земная кора просто не сохранила столь древних пород. К тому же ранние этапы пути могли быть пройдены и вовсе не на Земле, а в протопланетном облаке или на других небесных телах. Что касается «переходных звеньев», то все они вымерли, не оставив после себя прямых потомков, за исключением одного-единственного — так называемого «последнего универсального общего предка» всех современных живых существ. Специалисты называют его Лукой (Last Universal Common Ancestor, LUCA). Лука был уже довольно сложным организмом, напоминающим бактерию.
Доказательства единого происхождения: у всех – ДНК, РНК и белки; единый генетический код; близкие системы репликации, транскрипции и трансляции.
Поэтому приходится довольствоваться разработкой гипотез о том, КАК мог быть пройден долгий путь от мертвой материи к живой клетке. Эти гипотезы поддаются экспериментальной проверке, но таким способом можно доказать только принципиальную возможность того или иного хода событий. Гораздо труднее доказать, что все на самом деле именно так и происходило.
Тем не менее даже в этой крайне трудной области исследователи в последние годы добились заметных успехов.
Этапы зарождения жизни:
· Абиогенный синтез простых органических соединений (нет проблем)
· Абиогенный синтез сложных органических соединений – «кирпичиков» жизни (основные проблемы почти решены)
· Появление репликаторов (РНК?) (проблем еще много, но они постепенно решаются)
· После этого все уже намного проще, потому что начинает работать «дарвиновский» эволюционный механизм (наследственность, изменчивость, отбор)
· Появление всего остального: универсального механизма точного синтеза полипептидов (генетического кода и механизма трансляции), ДНК, липидных мембран и первых клеток.
· LUCA – Last Universal Common Ancestor

Наши предки и не подозревали, что вопрос о происхождении живого из неживого может стать серьезной проблемой для их потомков. Для них все вокруг было живым, одухотворенным. Мудрецы Античности и Средневековья не видели непреодолимой грани между живым и неживым. Вслед за Аристотелем вплоть до XVII века все ученые считали зарождение жизни самым обычным, повседневным явлением. В гниющих отходах зарождаются черви и мухи, в старом тряпье — мыши, на подводных камнях и днищах кораблей — моллюски. Могучая «животворная сила» пронизывает мироздание; она-то и заставляет косную материю порождать жизнь. Это учение — витализм — не противоречило и библейской версии космогенеза. «И сказал Бог: да произрастит земля зелень...» «И сказал Бог: да произведет вода пресмыкающихся...» Бог дал стихиям творческие силы. И с тех пор — чему же удивляться? — вода производила, а земля произращивала.
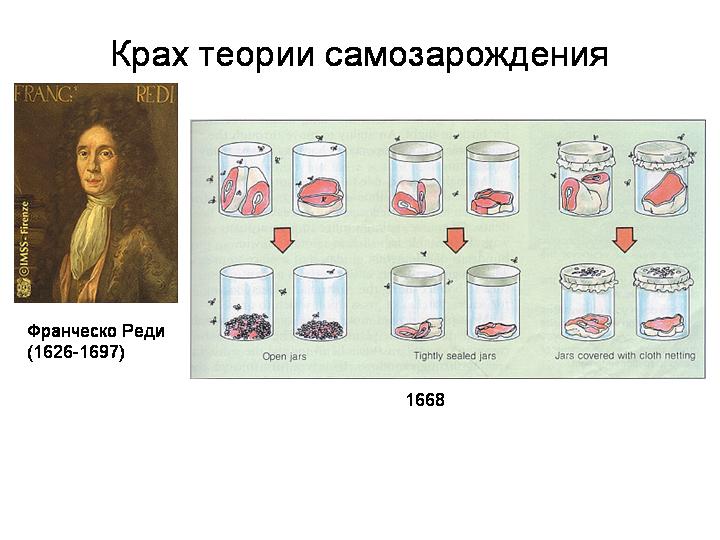
Реди доказал, что личинки мух не заведутся в тухлом мясе, если мясо накрыть тряпочкой, сделав его недоступным для взрослых мух.
Публика согласилась с Реди, что такие сложные животные, как мухи и мыши, могут рождаться только от соответствующих родителей. Но вот простые одноклеточные организмы — они-то, несомненно, порождаются грязной водой, гниющими отходами и тому подобным. Эта концепция просуществовала в общественном сознании около ста лет, а затем ученый монах Ладзаро Спалланцани обнаружил, что микробы тоже размножаются — делятся пополам, давая начало таким же микроорганизмам. Мало того, когда он кипятил бульон и плотно закрывал его крышкой, никаких микробов там не появлялось.

Однако многие ученые считали, что для зарождения жизни необходима «жизненная сила». Она не может проникнуть в закрытую банку, именно потому там и не появляются микробы. Луи Пастер в 1865 году хитроумным экспериментом сумел доказать, что никакой жизненной силы нет.
С витализмом было покончено, но все дивиденды достались не науке, а... церкви. Нелегкая задача встала перед теми учеными, которые не хотели привлекать сверхъестественные силы для объяснения природных явлений. После того как была доказана невозможность самозарождения, они должны были теперь долго и мучительно доказывать его возможность. Пусть не здесь и сейчас, а очень давно, и не за час-другой, а за миллионы лет.


Поначалу дело казалось безнадежным, ведь химики еще не умели получать органические вещества из неорганических. Грань между живой и неживой материей казалась непреодолимой. Правда, еще в 1828 г. Фридрих Велер синтезировал мочевину, но это никого не убедило. Однако в 1854 г. Марселен Бертло сумел искусственно получить липиды, а в 1864 г. А. М. Бутлеров открыл реакцию синтеза углеводов из формальдегида. Вскоре химики научились получать и другие органические вещества из неорганических.
Стало ясно, что между живой и неживой материей на химическом уровне нет непреодолимой грани. Это был первый крупный успех теории абиогенеза. Ученые поняли, что, хотя прямое самозарождение живых существ невозможно, жизнь могла появиться постепенно в результате долгой химической эволюции. Эта теория, в отличие от креационизма, является научной, то есть допускающей принципиальную возможность опровержения. Например, если бы оказалось, что органические вещества действительно невозможно получить из неорганических, как думали многие вплоть до опытов Бертло и Бутлерова, то абиогенез пришлось бы признать несостоятельной теорией. Однако этого не происходит. Наоборот, по мере развития науки один за другим проясняются все новые этапы долгого пути от неживой материи к первой клетке.
Дата добавления: 2015-06-10; просмотров: 2010;