ВИДООБРАЗОВАНИЕ
· Процесс, пограничный между «микроэволюцией» и «макроэволюцией».
· Вид – понятие довольно условное (см. в книге Дарвина). Биоразнообразие структурировано, но часто границы между «кластерами» размыты.
· Концепции вида: морфологическая (сходство/различие строения), биологическая (репродуктивная совместимость/ несовместимость), генеалогическая (обособленная ветвь на эволюционном дереве), экологическая (обособленная ниша, приспособленность к разным условиям).
· Многие виды являются таковыми по одним концепциям, но не являются – по другим.

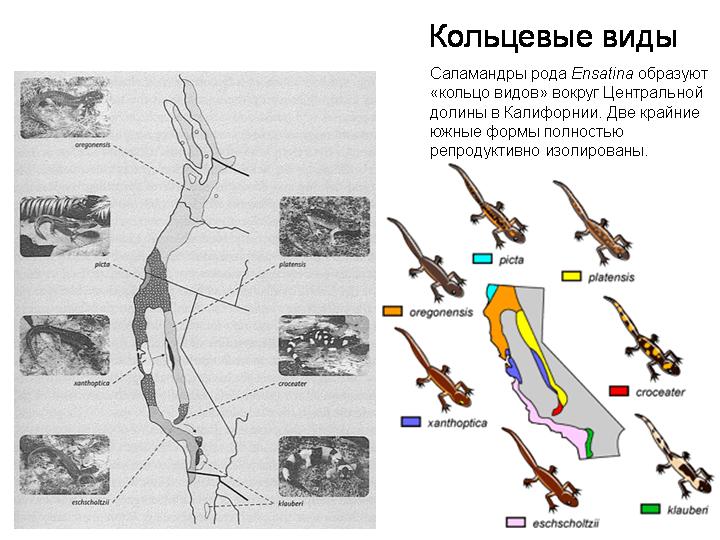
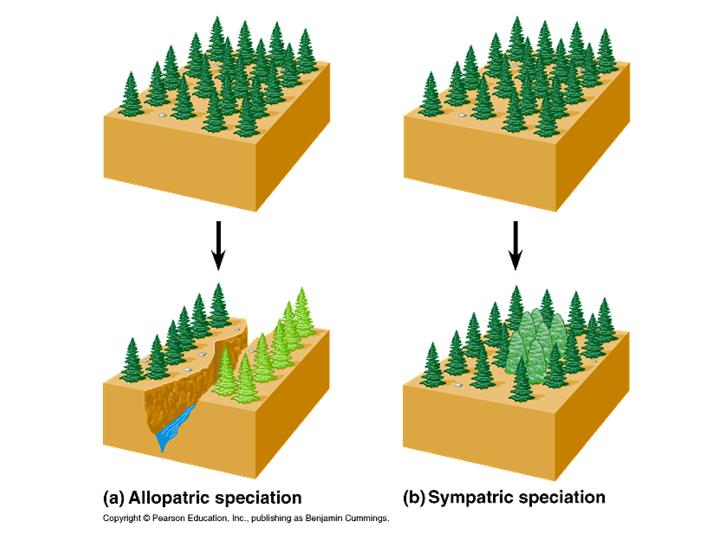
Ключевым моментом видообразования является формирование репродуктивной изоляции. Чтобы один исходный вид разделился на два, между двумя частями предкового вида должен прекратиться обмен генами. Если они продолжают активно скрещиваться, вид не сможет разделиться.
В роли первичного изолирующего фактора может выступать географический барьер (например, морской пролив или горный хребет). В этом случае говорят об аллопатрическом видообразовании. Барьер может быть экологическим или поведенческим (например, популяции могут «разойтись» по разным кормовым растениям, встречающимся на одной и той же территории); тогда видообразование называют симпатрическим.

Независимо от природы первичного изолирующего барьера, две разделившиеся популяции рано или поздно становятся генетически несовместимыми. Возникает т. н. постзиготическая изоляция, проявляющаяся в снижении жизнеспособности или плодовитости гибридного потомства.
Теперь, если две популяции снова встретятся и начнут скрещиваться, отбор будет поддерживать любые мутации, снижающие вероятность гибридизации и способствующие выбору «своих» в качестве брачных партнеров. Потому что скрещиваться с чужими просто-напросто невыгодно: потомство получится маложизнеспособным. Этот эффект называется «усилением» (reinforcement), и он приводит к тому, что представители двух разошедшихся популяций перестают даже пытаться спариться друг с другом. На этом последнем этапе могут быстро развиться самые разнообразные изолирующие механизмы, в том числе морфологические (например, различия в строении копулятивных органов) и поведенческие (разные брачные наряды, ритуалы). Эти механизмы уже можно считать настоящими адаптациями «для предотвращения гибридизации», поскольку они развились под действием отбора. И это уже будет не постзиготическая, а презиготическая изоляция.
Что же касается постзиготической изоляции, то она может возникать автоматически, сама собой, без помощи какого-либо «отбора на несовместимость» или других специальных механизмов – как неизбежный побочный эффект длительной разобщенности. Единственное, что для этого нужно, — это фактическая разобщенность двух популяций (отсутствие генетического обмена — неважно, по какой причине) и время. Об этом очень здорово написал в своей книге Дарвин – это одна из его гениальных догадок.
Времени может потребоваться много: до 10 млн лет или даже больше. В среднем – порядка 3 миллионов лет (для формирования полной постзиготической несовместимости). Если же на популяции действует интенсивный разнонаправленный отбор, постзиготическая изоляция может развиться очень быстро, всего за несколько сотен поколений.

В конце 30-х — начале 40-х годов прошлого века Добржанский и Мёллер предложили простую модель, согласно которой постзиготическая несовместимость разобщенных популяций развивается как неизбежный побочный эффект их разобщенности.
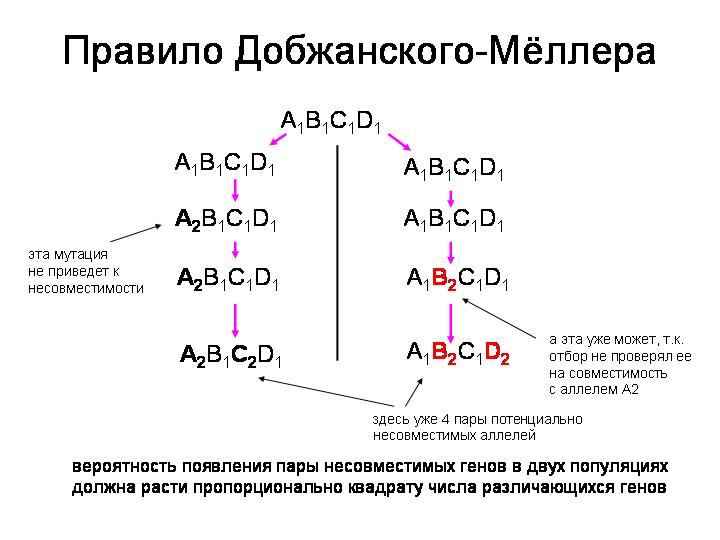
Допустим, в исходной популяции, еще до разделения, все особи имели генотип A1B1С1D1 (предположим для простоты, что организмы гаплоидные). Но вот популяция по какой-то причине разделилась на две изолированные части — например, несколько представителей исходной популяции занесло бурей на необитаемый остров. После этого в каждой из двух популяций будут независимо закрепляться разные мутации — неважно, под действием отбора или случайного дрейфа. Например, в популяции 1 закрепится мутация в локусе A. Теперь генотип у представителей этой популяции будет A2B1С1D1. Может ли это привести к постзиготической несовместимости с популяцией 2, где пока еще сохранился исходный генотип? Вряд ли, потому что ген A2, очевидно, вполне совместим с генами B1, C1 и D1 (иначе он бы не смог закрепиться в популяции 1). Но вот по прошествии какого-то времени в популяции 2 тоже закрепилась какая-нибудь мутация, например в локусе B. Теперь у них генотип A1B2С1D1. Ген B2, очевидно, совместим с A1, C1 и D1: это было «проверено» отбором, который в противном случае отбраковал бы ген B2 и не дал ему закрепиться в популяции 2. Однако ген B2 не проверялся отбором на совместимость с геном A2, который закрепился в популяции 1. Следовательно, с некоторой вероятностью эти гены могут оказаться несовместимыми.
Нетрудно заметить, что вероятность появления пары несовместимых генов в двух популяциях будет расти пропорционально квадрату числа различающихся генов (то есть квадрату «генетического расстояния» между популяциями). Например, если в каждой популяции закрепилось по одной мутации, то существует только одна пара потенциально несовместимых (не проверявшихся отбором на совместимость) генов. Если закрепилось по две мутации, то таких непроверенных отбором пар будет уже четыре: конфликт может возникнуть между A2 и B2, A2 и D2, C2 и B2, C2 и D2. Если в каждой популяции закрепилось по три мутации (в разных локусах), число потенциально несовместимых пар будет равно девяти, и так далее.
Таким образом, модель предсказывает, что число генов, ответственных за генетическую несовместимость, должно расти с ускорением, как снежный ком. Если предположить, что генетические различия накапливаются с постоянной скоростью, то число «генов несовместимости» должно быть пропорционально квадрату времени, прошедшего с момента разделения популяций.
В течение 70 с лишним лет, прошедших с момента появления этой модели, в ее работоспособности мало кто сомневался. Ее преподавали в университетах, а генетики, опираясь на нее, занимались поиском конкретных «генов несовместимости», порой успешно. Но вот проверить ее напрямую удалось только в 2010 году – причем сразу на двух объектах, на растениях и дрозофилах.
Drosophila melanogaster, D. simulans и D. santomea. Эволюционные пути видов mel и sim разошлись 5,4 млн лет назад, видов mel и san — 12,8 млн лет назад. Таким образом, время независимого существования в двух парах видов различается в 2,4 раза, и количество генетических различий тоже. Если гипотеза Добжанского–Мёллера верна, число «генов несовместимости» в паре mel-san должно быть примерно в 2,42 ≈ 6 раз больше, чем в паре mel-sim. Оказалось – в 5,74 раза (!).
И тоже самое получилось при анализе 4 видов рода Solanum (помидор и его дикие родственники). Число генов несовместимости действительно растет пропорционально квадрату генетических различий.
ДЗ: Прочесть раздел «Эмбриологические доказательства».
ВИДООБРАЗОВАНИЕ. Механизм «усиления» подтвержден эмпирически – например, на бабочках. У вторично-симпатрических видов различия по окраске максимальны, если эти виды разошлись недавно и еще сохранили (потенциально) способность к гибридизации. Если виды разошлись давно, они уже в принципе не могут скрещиваться, и у них менее контрастные различия по окраске.

До недавних пор вообще думали, что симпатрического видообразования не бывает. Симпатрическое видообразование было «не в чести» во многом из-за авторитета Эрнста Майра (1904-2005). Это была большая ошибка. Сейчас уже имеется много доказанных случаев.
Вообще-то ясно, что для появления каждого из многих миллионов видов обитающих на планете организмов физических барьеров просто не хватило бы — биосфера недостаточно сегрегирована для этого. Симпатрическое видообразование должно существовать, но было непонятно, каким образом оно может происходить. Предположим, что часть особей в популяции стала чем-то немного отличаться от остальных. Если эти особи не отделены от прочих физическим барьером, то любое отличие, казалось бы, должно быстро «размыться» в результате скрещиваний с неизменившимися сородичами. Полезное отличие станет общим достоянием, вредное исчезнет. В любом случае, вид не разделится на два, если у «немного отличающихся» особей не возникнет эндогамия, т. е. предпочтение себе подобных в качестве брачных партнеров. Как может вознинкуть эндогамия, было не совсем понятно.

В 2006 году был описан случай видообразования, в симпатрическом характере которого трудно усомниться. Речь идет о двух видах рыб — цихлид, живущих в маленьком озере Апойо в Никарагуа. Это круглое озеро диаметром 5 км и глубиной до 200 м представляет собой залитый водой вулканический кратер. Озеро полностью изолировано от других водоемов и образовалось менее 23 тыс. лет назад.
В озере живет два вида цихлид: широко распространенный Amphilophus citrinellus и встречающийся только в этом озере A. zaliosus.
Сравнили последовательности митохондриальной ДНК у 120 рыб обоих видов из оз. Апойо и более 500 представителей A. citrinellus из других озер. Анализ показал, что все рыбы из Апойо, относящиеся к обоим видам, образуют монофилетическую ветвь (т. е. имеют единое происхождение), причем варианты мтДНК из оз. Апойо не встречаются в других озерах. Попросту говоря, это означает, что озеро было заселено единожды, повторных колонизаций и обмена генами с жителями других озер не было, и все живущие ныне в озере рыбы, вероятно, являются потомками одной самки, когда-то попавшей в озеро. И самка эта относилась к виду A. citrinellus.
Анализ ядерных генов подтвердил результаты, основанные на мтДНК.
Статистический анализ всей совокупности данных по генетическому разнообразию цихлид из озера Апойо показал, что два вида полностью репродуктивно изолированы (не скрещиваются друг с другом). О том же свидетельствуют и поведенческие эксперименты, показавшие, что эти рыбы безошибочно выбирают себе в качестве брачных партнеров представителей своего вида.
Эти рыбы и внешне достаточно хорошо различаются (т.е. по морфологическому критерию это тоже разные виды). Образ жизни у них тоже разный. A. citrinellus держится вблизи дна, более всеяден, не брезгует крупными водорослями (харовыми) и редко глотает насекомых; A. zaliosus плавает в толще воды, не ест водорослей, зато насекомых очень любит. Всё это говорит о том, что мы имеем дело с бесспорным случаем симпатрического видообразования.
Дата добавления: 2015-06-10; просмотров: 1312;