Значение фотосинтеза. Лист как орган фотосинтеза
1. Значение процесса фотосинтеза и история его изучения
Анализ органического вещества показывает, что оно состоит на 45% из углерода. Именно поэтому вопрос об источнике питания организмов углеродом чрезвычайно важен. Все организмы можно разделить на автотрофные и гетеротрофные. Автотрофные организмы характеризуются способностью синтезировать органическое вещество из неорганических соединений. Гетеротрофные организмы строят органическое вещество своего тела из уже имеющихся готовых органических соединений, только перестраивая их. Иначе говоря, гетеротрофные организмы живут за счет автотрофных. Для того чтобы осуществить синтез органического вещества, необходима энергия.
В зависимости от используемого соединения, а также от источников энергии, различают следующие основные типы питания углеродом и построения органических веществ (таблица 1).
Таблица 1 - Основные типы углеродного питания
| Тип питания | Источник углерода | Источник водорода | Источник энергии |
| Гетеротрофный | Органическое вещество | Органическое вещество | Энергия окисления органических веществ |
| Автотрофный 1. Фотосинтез | СО2 | Н2О | Энергия света |
| 2.Бактериальный фотосинтез | СО2 | Н2О, Н2 и др. | Энергия света |
| 3. Хемосинтез | СО2 | Н2О, H2S, H2,NH3 | Энергия окисления неорганических веществ |
Из всех перечисленных типов питания углеродом фотосинтез зеленых растений, при котором построение органических соединений идет за счет простых неорганических веществ (С02 и Н2О) с использованием энергии солнечного света, занимает совершенно особое место. Общее уравнение фотосинтеза:
6СО2 + 12Н2О → С6Н12О6 + 6О2 + 6Н2О
Фотосинтез— это процесс, при котором энергия солнечного света превращается в химическую энергию. В самом общем виде это можно представить следующим образом: квант света (hv) поглощается хлорофиллом, молекула которого переходит в возбужденное состояние, при этом электрон переходит на более высокий энергетический уровень. В клетках зеленых растении в процессе эволюции выработался механизм, при котором энергия электрона, возвращающегося на основной энергетический уровень, превращается в химическую энергию.
Только с помощью зелёных растений энергия Солнца может накапливаться в виде энергии химических связей. Большая часть энергии, используемой человеком на заводах и фабриках, т. е. энергия, благодаря которой происходит движение различных механизмов, машин и самолетов,— это все энергия Солнца, преобразованная в зеленом листе. Запасание энергии в результате фотосинтеза происходит на различные промежутки времени: от минут, часов до сотен миллионов лет (достаточно вспомнить образование торфа и каменного угля в результате разложения растений).
В процессе фотосинтеза из простых неорганических соединений (СО2, Н-О) строятся, различные органические вещества. В результате происходит перестройка "химических связей: вместо связей С—О и Н—О возникают связи С—С и С—Н, в которых электроны занимают более высокий энергетический уровень. Таким образом, богатые энергией органические вещества, которыми питаются и за счет которых получают энергию (в процессе дыхания) животные и человек, первоначально создаются в зеленом листе. Можно сказать, что практически вся живая материя на Земле является результатом фотосинтетической деятельности. Исследования показали также, что почти весь кислород атмосферы фотосинтетического происхождения. Следовательно, процессы дыхания и горения стали возможны только после того, как возник фотосинтез. Все это и позволяет говорить о космическом значении фотосинтеза. Появление свободного кислорода в атмосфере Земли вызвало значительные изменения во всей живой природе. Возникли аэробные организмы, способные усваивать кислород. На поверхности Земли процессы приняли биогеохимический характер, произошло окисление соединений железа, серы, марганца и др. Изменился состав атмосферы: содержание СО2 и аммиака снизилось, а кислорода и азота возросло. Возникновение озонового экрана, который задерживает опасную для живых организмов ультрафиолетовую радиацию, также является следствием появления кислорода. Озон (О3) образуется из О2 в верхних слоях атмосферы под действием солнечной радиации. В настоящее время существует опасность частичного разрушения озонового экрана вследствие загрязнения атмосферы промышленными и другими отходами.
Фотосинтез имеет важнейшее значение и в жизни самого растительного организма, являясь процессом воздушного питания растений, Согласно расчетам А.А. Ничипоровича, в период наиболее активного роста растений суточные приросты сухого вещества достигают 300 и даже 500 кг/га. При этом в течение суток растение усваивает 1—2 кг N, 0,1—0,2 кг Р, 0,8—1,7 кг К и до 1000 кг СО2. Чтобы лучше представить себе масштабы процесса фотосинтеза, приведем несколько цифр. Согласно данным французского исследователя Дювиньо (1972), ежегодно в процессе фотосинтеза растениями суши образуется 30 млрд т органического вещества, в том числе на долю лесов приходится 20,4 млрд т, лугов, степей — 3 млрд т, пустынь — 1,1 млрд т, культурных полей — 5,6 млрд т. Энергия, накапливаемая в процессе фотосинтеза за один год, приблизительно в 100 раз больше энергии сгорания всего добываемого в мире за этот же период угля. Казалось бы, что при таком колоссальном годовом потреблении углерода весь СО2 воздуха должен быть израсходован в течение немногих лет. Однако содержание СО2 в атмосфере Непрерывно пополняется за счет растворенных в воде карбонатов и бикарбонатов. Кроме того, углекислый газ выделяется из почвы в результате различных микробиологических процессов, связанных с окислением органических веществ (до 25—30 кг СО2 на 1 га в сутки) и др. Учитывая значение процесса фотосинтеза, раскрытие его механизма является одной из наиболее важных и интересных задач, стоящих перед физиологией растений.
Датой открытия процесса фотосинтеза можно считать 1771 г. Английский ученый Дж. Пристли обратил внимание на изменение состава воздуха вслёдствие жизнедеятельности животных. В присутствии зеленых растений воздух вновь становился пригодным как для дыхания, так и для горения. В дальнейшем работами ряда ученых (Я. Ингенгауз, Ж. Сенебьё, Т. Соссюр, Ж. Б. Буссенго) было установлено, что зеленые растения из воздуха поглощают СО2, из которого при участии воды на свету образуется органическое вещество. Именно этот процесс в 1877 немецкий ученый В. Пфеффер назвал фотосинтезом. Большое значение для раскрытия сущности фотосинтеза имел закон  сохранения энергии сформулированный Р. Майером. В 1845 г. Р. Майёр выдвинул предположение, что энергия, используемая растениями, — это энергия Солнца, которою растения в процессе фотосинтеза превращают в химическую энергию. Это положение было развито и экспериментально подтверждено в исследованиях замечательного русского ученого К.А. Тимирязева.
сохранения энергии сформулированный Р. Майером. В 1845 г. Р. Майёр выдвинул предположение, что энергия, используемая растениями, — это энергия Солнца, которою растения в процессе фотосинтеза превращают в химическую энергию. Это положение было развито и экспериментально подтверждено в исследованиях замечательного русского ученого К.А. Тимирязева.
2. Лист как орган фотосинтеза. Особенности диффузии СО2 в листе
Фотосинтез может протекать в различных органах растений (стебли, плоды и др.), имеющих зеленую окраску, но основным органом фотосинтеза является лист. Анатомическое строение листа приспособлено к тому, чтобы обеспечить поступление СО2 к клеткам, содержащим зеленые пластиды, и достигнуть максимального поглощения энергии света. Листья в большинстве случаев тонкие и обладают максимальной поверхностью на единицу массы. Наличие межклетников облегчает доступ СО2 ко всем клеткам. К эпидермису, расположенному на верхней стороне листа, примыкает палисадная паренхима, клетки которой вытянуты перпендикулярно поверхности листа. Палисадная паренхима — это основная ассимиляционная ткань листа, особенно богатая хлоропластами. Густая сеть жилок в листе не только облегчает снабжение клеток паренхимы водой, но и способствует быстрому оттоку из листа углеводов, образующихся в процессе фотосинтеза.
Для того чтобы процесс фотосинтеза протекал нормально, в клетки к зеленым пластидам должен непрерывно поступать СО2. Основным поставщиком СО2 служит атмосфера. Количество СО2 в атмосфере составляет около 0,03%. В течение дня растения усваивают количество СО2, содержащееся в 30—60-метровом слое воздуха. Для образования 1 г сахара необходимо 1,47 г СО2, содержащегося в 2500 л воздуха. Это возможно благодаря непрерывному турбулентному движению воздушных масс вокруг листьев, вызываемому ветром и неравномерным нагреванием их солнечными лучами. Углекислый газ, потребляемый при фотосинтезе, возвращается в атмосферу за счет процессов дыхания и гниения. Особенное значение имеет при этом деятельность почвенных микроорганизмов. Определенное количество СО2 выделяется при разложении карбонатов, растворенных в морской воде. В результате поглощения листом СО, возникает градиент концентрации этого газа, что и вызывает непрерывную диффузию СО2 в направлении фотосинтезирующих органов растения. Диффузия возникает вследствие хаотического теплового движения молекул и представляет спонтанный процесс, приводящий к перемещению вещества от его большей концентрации к меньшей. Согласно закону Фика скорость диффузии прямо пропорциональна разности концентраций и обратно пропорциональна сопротивлению. СО2 диффундирует из более дальних слоев атмосферы в близлежащие к листу и далее в межклетные пространства, из межклетников в клетки и далее к хлоропластам. Чем быстрее используется СО2 в процессе фотосинтеза, тем больше падает ее парциальное давление в межклетниках и тем быстрее поступает в них СО2. Всякое перемешивание среды (воздуха или воды) способствует более быстрой диффузии СО2 к листу. В процессе диффузии ток СО2 встречает сопротивление. Оно особенно велико при диффузии СО2 к листьям водных растений. Исследования показали, что в воде сопротивление диффузионному току СО2 примерно в 1000 раз больше, чем в воздухе. Кроме внешнего сопротивления, которое встречает СО2 при диффузии до поверхности листа, существует еще внутреннее сопротивление (в самом листе).
Углекислый газ поступает в лист растения через устьица. Некоторое количество СО2 поступает непосредственно через кутикулу. В последнем случае диффузия СО2 происходит через клетки эпидермиса к хлоропластам клеток паренхимы листа. При прохождении через устьичные щели СО2 может диффундировать в виде газа к любой части листа по межклеточным пространствам. Расстояние, которое должны преодолеть молекулы СО2 по системе межклетников до клетки, составляет около 1000 мкм. Время прохождения этого расстояния — 10—16 микросекунд. В этом случае водный диффузионный путь минимальный — лишь внутри клетки, а следовательно, сопротивление будет меньшим. Несмотря на то, что при полностью открытых устьицах площадь устьичных щелей составляет всего 1/100 поверхности листа, диффузия СО2 внутрь листа идет через них сравнительно быстро. Опытным путем установлено, что свободная поверхность щелочи площадью 1 см2 поглощает за 1 ч 0,12—0,15 см3 СО2. 1 см2 поверхности листа поглощает всего в два раза меньше — 0,07 см3 СО2, в то же время его открытая площадь меньше в 100 раз. Такая высокая скорость связана с тем, что диффузия газов через мелкие отверстия идет пропорционально не их площади, а диаметру. Естественно, что это положение правильно лишь при условии, что устьица открыты. При закрытых устьицах диффузия СО2 в лист резко сокращается. При ветре внешнее сопротивление падает. Основное значение приобретает сопротивление при диффузии через устьица, поэтому их закрытие оказывает еще большее относительное влияние и еще сильнее снижает диффузию СО2.
Закрытие устьиц сильнее уменьшает потерю паров воды из листа (транспирацию) по сравнению с диффузией СО2 внутрь листа. Это связано с тем, что скорость диффузии паров воды в первую очередь зависит от размера устьичных щелей, тогда как для диффузии СО2 большее значение имеет интенсивность его использования в процессе фотосинтеза. Это представляет значительное приспособление для выживания растений в неблагоприятных условиях существования, например при засухе. Так, уменьшение диаметра устьиц с 10 до 3 мкм уменьшает транспирацию на 38%, тогда как поглощение СО2 — всего на 29%.
3. Хлоропласты, их строение и образование
Весь процесс фотосинтеза протекает в зеленых пластидах — хлоропластах. Различают три вида пластид: лейкопласты — бесцветные, хромопласты — оранжевые, хлоропласты — зеленые. В лейкопластах синтезируется и отлагается в запас крахмал, в хромопластах накапливаются каротиноиды, в хлоропластах сосредоточен зеленый пигмент хлорофилл и происходит фотосинтез.
Незеленые организмы, например грибы, лишены пластид. Эти растения не обладают способностью к фотосинтезу. В процессе эволюции дифференциация пластид произошла очень рано. Правда, у фотосинтезирующих бактерий пластид еще нет, их роль выполняют внутрицитоплазматические мембраны (пурпурные бактерии) или особые структуры — хлоросомы, локализованные на мембранах (зеленые бактерии). Это наиболее примитивная организация фотосинтетического аппарата. Однако уже у водорослей имеются специальные образования (хроматофоры), в которых сосредоточены пигменты, они разнообразны по форме (спиральные, ленточные, в виде пластинок или звезд). Высшие растения характеризуются вполне сформировавшимся типом пластид в форме диска или двояковыпуклой линзы. Приняв форму диска, хлоропласты становятся универсальным аппаратом фотосинтеза.
Размер хлоропластов колеблется от 4 до 10 мкм. Число хлоропластов обычно составляет от 20 до 100 на клетку. Химический состав хлоропластов достаточно сложён и может быть охарактеризован следующими средними данными (% на сухую массу): белок —35—55; липиды — 20—30; углеводы — 10; РНК — 2—3; ДНК — до 0,5; хлорофилл — 9; каротиноиды — 4,5.
Важно отметить, что многие белки хлоропластов обладают ферментативной активностью. Действительно, в хлоропластах сосредоточены все ферменты принимающие, участие в процессе фотосинтеза (окислительно-восстановительные, синтетазы. гидролазы). В настоящее время доказано, что в хлоропластах, так же как и в митохондриях, имеется своя белоксинтезирующая система. Многие из ферментов, локализованных в хлоропластах, являются двухкомпонентными. Во многих случаях простетическая группа ферментов — это различные витамины. В хлоропластах сосредоточены многие витамины и их производные (витамины группы В, К, Е, D). В хлоропластах находится 80% Fe, 70% Zn, около 50% Си от всего количества этих элементов в листе.
Внутреннее строение хлоропластов, их ультраструктура были раскрыты с использованием электронного микроскопа, Оказалось, что хлоропласты окружены двойной мембраной. Толщина каждой мембраны 7,5—10 нм, расстояние между ними — 30 нм. Внутреннее пространство хлоропластов заполнено бесцветным содержимым — стромой и пронизано мембранами (ламеллами). Ламеллы, соединенные друг с другом, образуют как бы пузырьки—тилакоиды (греч. «тилакоидес» — мешковидный). В хлоропластах содержатся тилакоиды двух типов. Короткие тилакоиды собраны в пачки и расположены друг над другом, напоминая стопочку монет. Эти стопки называются гранами, а составляющие их тилакоиды — тилакоидами гран. Между гранами параллельно друг другу располагаются длинные_тилакоиды. Составляющие их ламеллы получили название — тилакоиды стромы. Между отдельными тилакоидами в стопках гран имеются узкие щели. Тилакоидные мембраны содержат большое количество белков, участвующих в фотосинтезе. В составе интегральных мембранных белков имеется много гидрофобных аминокислот. Это создает безводную среду и делает мембраны стабильнее. Многие белки тилакоидных мембран построены в виде векторов играничат с одной стороны со стромой, а с другой контактируют с внутренним пространством тилакоида.
Относительно связи между ламеллами гран и ламеллами стромы имеются разные точки зрения. Т .Вейер предложил гранулярно-решетчатую модель, согласно которой внутренние пространства всех тилакоидов соединены между собой (рис.1).Таким образом, в хлоропластах имеется как бы два раздельных пространства — внутреннее (внутри тилакоидов) и внешнее (вне тилакоидов). У большинства водорослей гран нет, а ламеллы собраны в группы (пачки) по 2—8 штук. Не во всех случаях и у высших растений хлоропласта имеют гранальную структуру. Так, в листьях кукурузы имеются два вида хлоропластов. В клетках мезофилла содержатся мелкие хлоропласты гранального строения. В клетках обкладки, окружающих листовые сосудистые пучки, хлоропласты крупные и гран не содержат.
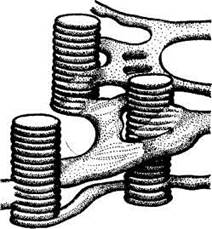
 Рисунок 1 - Схема гранулярно-сетчатой структуры хлоропластов
Рисунок 1 - Схема гранулярно-сетчатой структуры хлоропластов
В строме хлоропластов находятся нити ДНК, рибосомы, крахмальные зерна. Основной фермент обеспечивающий усвоение углекислого газа, — рибузолобифосфаткарбоксилазаоксигеназа (сокращенно РБФ — карбоксилаза оксигеназа) также расположен в строме. Зеленый пигмент хлорофилл в виде комплекса с белками (пигмент-белковые комплексы) сосредоточен главным образом в тилакоидах гран и частично в тилакоидах стромы. В мембранах тилакоидов гран локализован фермент, катализирующий синтез АТФ (АТФ-синтаза). Этот фермент связан с белком, расположенным в самой мембране (рис. 2).
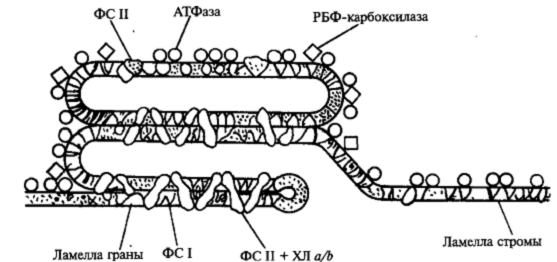
Рисунок 2 – Поверхность ламелл хлоропластов
4.Онтогенез пластид
Пластиды, так же как и митохондрии, не возникают вновь, а размножаются путем деления. В яйцеклетке имеются так называемые инициальные частицы, из которых в дальнейшем и развиваются как митохондрии, так и пластиды. Эта точка зрения подтверждается явлением так называемой цитоплазматической или пластидной (внехромосомной) наследственности. Как известно, у раздельнополых организмов женские и мужские гаметы вносят в зиготу одинаковый вклад в отношении генов. Однако женская гамета содержит во много раз больше цитоплазмы и инициальных частиц. Корренс показал, что окраска листьев у пестролистных растений наследуется исключительно по материнской линии. Так, оказалось, что цветки, развившиеся на зеленых побегах, дают семена, из которых вырастают растения с зелеными листьями. Цветки на ветвях с пестрыми листьями дают семена, из которых вырастают пестролистные растения. Окраска листьев растений, с цветков которых собиралась пыльца для опыления, не имеет значения для потомства. Эти опыты и привели к предположению, что хлоропласты представляют собой генетически автономные образования и их свойства наследуются по материнской линии.
Так же как и для митохондрии, начальной стадией роста хлоропластов являются инициальные частицы (рис. 3).Эти частицы — глобулярные образования, окруженные двойной мембраной значительно более плотной консистенции по сравнению с окружающей гиалоплазмой. Инициальные частицы увеличиваются в размере и приобретают форму двояковыпуклой линзы. Одновременно их внутренняя мембрана начинает разрастаться, образуя складки. От складок отшнуровываются пузырьки (тилакоиды), которые располагаются параллельно и пронизывают всю строму. На этой стадии развития частицы становятся видимыми в световой микроскоп (0,3—0,5 мкм) — это уже пропластиды.
Для дальнейшего развития структуры пропластид необходим свет. На свету образуется хлорофилл. Молекулы хлорофилла локализуются в мембранах. Именно на свету образуются два типа тилакоидов. Длинные тилакоиды тянутся через все продольное сечение пластид и образуют тилакоиды стромы. Короткие тилакоиды располагаются стопкой друг над другом и образуют тилакоиды гран.
Пластиды достигают окончательного размера. Непосредственно из пропластид могут образовываться и бесцветные пластиды (лейкопласты-амилопласты). Лейкопластычаще всего локализованы в клетках запасающих тканей. Подобно пропластидам они характеризуются слабо развитой ламеллярной структурой. Во многих случаях в лейкопластах ламеллы сохраняют связь с внутренней оболочкой. В строме лейкопластов располагаются крахмальные зерна, осмиофильные глобулы, белковые включения. Амилопласты могут превращаться в хлоропласты, например, как это происходит при позеленении клубней картофеля на свету. Хромопласты— это, по-видимому, результат деградации хлоропластов, при которой ламеллярная структура частично разрушается. Одновременно происходит образование осмиофильных глобул, содержащих каротиноиды. Эти глобулы располагаются сплошным слоем под оболочкой пластид.
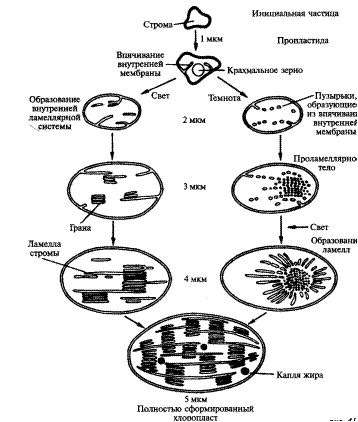
Рисунок 4 – Онтогенез пластид
В клетках растущих листьев пластиды размножаются путем деления. В листьях шпината при увеличении листа с 1 см до полного размера количество пластид на клетку возрастает с 50 до 500 штук. Процесс деления пластид ускоряется на свету. Особенное значение имеют красные и синие участки спектра, которые поглощаются хлорофиллом. Деление может происходить или на стадии пропластид (у высших растений), или на стадии уже сформировавшихся хлоропластов (у водорослей и папоротников). Пластиды делятся или путем возникновения поперечных перегородок, или, реже, путем почкования. Поперечные перегородки образуются за счет образования складок на внутренней мембране. До тех пор, пока складка внутренней мембраны не поделила тело пластид, наружная мембрана сохраняется и объединяет обе дочерние пластиды. Дальнейшее поведение наружной мембраны неясно. Она либо разрывается, либо образует складки. Таким образом, пластиды — самоудваивающиеся органеллы.
5. Полуавтономность хлоропластов и их происхождение
Хлоропласты содержат специфические молекулы ДНК и обладают белоксинтезирующей системой. Однако хлоропласты нельзя отнести к полностью автономным образованиям. В самом деле, количество находящейся в пластидах ДНК не может обеспечить все разнообразие пластидных белков. Частично белки хлоропластов образуются под контролем ядра (Ю.С. Насыров).
Хлоропластная ДНК представляет собой кольцевую двуспиральную молекулу и отличается от линейных молекул ДНК в хромосомах ядра. В ней зашифрована информация о рРНК и тРНК хлоропластов, 50% белков тилакоидной мембраны, рибосомальных белков, а также ферменте РНК-полимеразе, который осуществляет синтез мРНК на хлоропластной ДНК. ДНК хлоропластов содержит гены больших субъединиц ключевого фермента фотосинтеза РБФ-карбоксилазы/оксигеназы. Остальные структурные белки и белки-ферменты кодируются в ядре. Собственный генетический аппарат имеют и митохондрии. Об этом уже говорилось выше. Геном хлоропластов меньше, чем митохондрий. Молодые митохондриальные клетки обычно полиплоидны и содержат много хромосом. Хлоропласты становятся полиплоидны только в зрелой клетке и при этом количество ДНК больше чем в митохондриях. В целом сумма генома хлоропластов и митохондрий составляет около трети генома ядра.
В организации генетического аппарата хлоропластов имеются сходные черты с генетическим аппаратом, как бактерий, так и эукариот. По прокариотическому типу организованы промоторы и терминаторы. Однако в ДНК хлоропластов обнаружены интроны, характерные для генов эукариот. Белоксинтезирующая система хлоропласта также имеет специфические черты и отличается от белоксинтезирующей системы цитоплазмы. Хлоропластные рибосомы мельче цитоплазматических и относятся к 70 S типу, свойственному для прокариот. В цитоплазме содержатся рибосомы 80 S, т. е. эукариотического типа. Как уже рассматривалось, синтез белка в хлоропластах и цитоплазме подавляется разными антибиотиками. Использование ингибиторов белкового синтеза позволило ученым точно установить, где в клетке (в хлоропласте или цитоплазме) синтезируется тот или иной белок.
Таким образом, полуавтономность хлоропласта проявляется в том, что его рост, деление и развитие находятся под контролем как генома ядра, так и хлоропласта. Интеграция работы двух геномов проявляется на уровне формирования сложных пептидных комплексов. Каждый такой комплекс состоит из белков, часть которых кодируется и синтезируется в хлоропласте, а часть кодируется в ядре, синтезируется на 80 S рибосомах цитоплазмы, а затем проникает в хлоропласт и участвует в построении комплексов. Например, большие субъединицы РБФ-карбоксилазы/оксигеназы кодируются и синтезируются в хлоропласте, а малые кодируются в ядре, синтезируются в цитоплазме в виде предшественника и проникают в хлоропласт. Для образования правильной структуры фермента необходим еще дополнительный белок — шаперон, который обеспечивает правильную укладку цепей субъединиц. Этот белок также кодируется в ядре и синтезируется в цитоплазме, проникая затем в хлоропласт. Установление зависимости формирования важнейших структур хлоропласта от ядра и цитоплазмы помогло объяснить невыполнимость идеи создания культуры изолированных хлоропластов, где бы они самостоятельно размножались на питательной среде.
Большой интерес имеет вопрос о возникновении хлоропластов в клетке в процессе эволюции. Поскольку хлоропласты представляют собой относительно независимое от ядра образование, способное к делению, росту, дифференциации, возникла гипотеза о том, что на заре эволюции хлоропласты, так же как и митохондрии, являлись самостоятельными организмами. Согласно этой симбиотрофной гипотезе хлоропласты возникли в результате симбиоза какого-то автотрофного организма (возможно, цианобактерии), способного трансформировать энергию солнечного света, с гетеротрофной клеткой. Пластиды и фотосинтезирующие прокариоты очень сходны между собой (кольцевая структура ДНК, размеры рибосом и др.). Согласно гипотезе, развиваемой рядом исследователей, способность к фотосинтезу возникла у прокариотов. Впоследствии гетеротрофные эукариотические клетки приобрели способность к фотосинтезу благодаря заглатыванию микробов-фотосинтетиков. Это событие (около 400 млн лет назад) и привело к возникновению фотосинтезирующих водорослей (Л. Маргелис, 1983). В этой связи интересно, что в 1969 г. было показано, что изолированные клетки млекопитающих способны заглатывать путем фагоцитоза выделенные из листьев хлоропласты. Захваченные клетками хлоропласты выживали на протяжении шести клеточных делений. При этом они сохраняли структуру и делились. Вновь выделенные из этих клеток хлоропласты не потеряли способности к фотосинтезу.
6. Физиологические особенности хлоропластов
Важным свойством хлоропластов является их способность к движению. Хлоропласты передвигаются не только вместе с цитоплазмой, но способны и самопроизвольно изменять свое положение в клетке. Скорость движения хлоропластов составляет около 0,12 мкм/с. Хлоропласты могут быть распределены в клетке равномерно, однако чаще они скапливаются около ядра и вблизи клеточных стенок. Большое значение для расположения хлоропластов в клетке имеют направление и интенсивность освещения. При малой интенсивности освещения хлоропласты становятся перпендикулярно к падающим лучам, что является приспособлением к лучшему их улавливанию. При высокой освещенности хлоропласты передвигаются к боковым стенкам и поворачиваются ребром к падающим лучам. Получены данные, что движение хлоропластов регулируется синим светом. В зависимости от освещения может также меняться и форма хлоропластов. При более высокой интенсивности света их форма становится ближе к сферической.
Основная функция хлоропластов — это фотосинтез. В 1955 г. Д. Арнон показал, что в изолированных хлоропластах может быть осуществлен весь процесс фотосинтеза. Важно отметить, что хлоропласты имеются не только в клетках листа. Они встречаются в клетках не специализирующихся на фотосинтезе органов: в стеблях, колосковых чешуйках и остях колосьев, корнеплодах, клубнях картофеля и т. д. В ряде случаев зеленые пластиды обнаруживаются в тканях, расположенных не в наружных, освещенных частях растений, а в слоях, удаленных от света: в тканях центрального цилиндра стебля, в средней части луковицы лилейных, а также в клетках зародыша семени многих покрытосеменных растений. Последнее явление (хлорофиллоносность зародыша) привлекает внимание систематиков растений. Имеются предложения разделить все покрытосеменные растения на две большие группы: хлороэмбриофиты и лейкоэмбриофиты, содержащие и не содержащие хлоропласты в зародыше (М.С. Яковлев).
Исследования показали, что структура хлоропластов, расположенных в других органах растения, так же как и состав пигментов, сходны с хлоропластами листа. Это дает основания считать, что они способны к фотосинтезу. В том случае, если они подвергаются освещению, по-видимому, в них действительно происходит фотосинтез. Так, фотосинтез хлоропластов, расположенных в остях колоса, может составлять около 30% от общего фотосинтеза растения. Позеленевшие на свету корни способны к фотосинтезу. В хлоропластах, находящихся в кожуре плода до определенного этапа его развития, также может идти фотосинтез. Согласно предположению А.Л. Курсанова, хлоропласты, расположенные вблизи проводящих путей, выделяя кислород, способствуют повышению интенсивности обмена веществ ситовидных трубок.
Вместе с тем роль хлоропластов не ограничивается их способностью к фотосинтезу. В определенных случаях они могут служить источником питательных веществ {Е.Р. Гюббенет). Хлоропласты содержат большое количество витаминов, ферментов и даже фитогормонов {в частности, гиббереллина и абсцизовой кислоты). В условиях, при которых ассимиляция исключена, зеленые пластиды могут играть активную роль в процессах обмена веществ. Известно, что присутствие листьев, находящихся в условиях, исключающих фотосинтез (закрывание землей), улучшает рост растений.
ФОТОСИНТЕЗ (12 часов)
Дата добавления: 2015-06-10; просмотров: 8259;