Закономерности независимого наследования двух и более признаков. Виды взаимодействия неаллельных генов.
Независимое наследование признаков. Такой характер наследования признаков впервые был описан Г. Менделем в опытах на горохе, когда одновременно анализировалось наследование в ряду поколений нескольких признаков, например цвета и формы горошин (рис. 6.11). Каждый из них в отдельности подчинялся закону расщепления в F2. В то же время разные варианты этих признаков свободно комбинировались у потомков, встречаясь как в сочетаниях, наблюдаемых у их родителей (желтый цвет и гладкая форма или зеленый цвет и морщинистая форма), так и в новых сочетаниях (желтый цвет и морщинистая форма или зеленый цвет и гладкая форма). На основании анализа полученных результатов Г. Мендель сформулировал закон независимого наследования признаков, в соответствии с которым: «Разные пары признаков, определяемые неаллельными генами, передаются потомкам независимо друг от друга и комбинируются у них во всех возможных сочетаниях».
Очевидно, этому закону должны подчиняться в первую очередь неаллельные гены, располагающиеся в разных (негомологичных) хромосомах. В таком случае независимый характер наследования признаков объясняется закономерностями поведения негомологичных хромосом в мейозе. Названные хромосомы образуют со своими гомологами разные пары, или биваленты, которые в метафазе I мейоза случайно выстраиваются в плоскости экватора веретена деления. Затем в анафазе I мейоза гомологи каждой пары расходятся к разным полюсам веретена независимо от других пар. В результате у каждого из полюсов возникают случайные сочетания отцовских и материнских хромосом в гаплоидном наборе (см. рис. 3.75). Следовательно, различные гаметы содержат разные комбинации отцовских и материнских аллелей неал-лельных генов.
Разнообразие типов гамет, образуемых организмом, определяется степенью его гетерозиготности и выражается формулой 2n, где n — число локусов в гетерозиготном состоянии. В связи с этим дигетерозиготные гибриды F1 образуют четыре типа гамет с одинаковой вероятностью. Реализация всех возможных встреч этих гамет при оплодотворении приводит к появлению в F2 четырех фенотипических групп потомков в соотношении 9:3:3:1. Анализ потомков F2 по каждой паре альтернативных признаков в отдельности выявляет расщепление в соотношении 3:1.
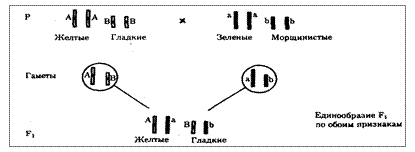
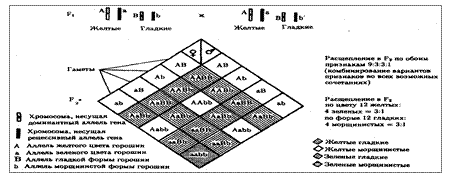
Рис. 6.11. Независимое наследование признаков (цвета и формы горошин)
Открытие независимого характера наследования разных признаков у гороха дало возможность Г. Менделю высказать предположение о дискретности наследственного материала, в котором за каждый признак отвечает своя пара наследственных задатков, сохраняющих в ряду поколений свою структуру и не смешивающихся друг с другом. Современные представления о надмолекулярной организации наследственного материала в хромосомах и закономерностях их передачи в ряду поколений клеток и организмов объясняют независимый характер наследования признаков расположением соответствующих генов в негомологичных хромосомах.
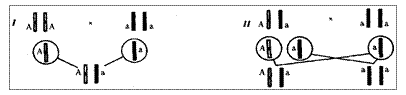
Рис. 6.12. Анализирующее (моногибридное) скрещивание
Объяснение см. в тексте
Наследование при комплементарном взаимодействии генов. Если сложный признак формируется в результате взаимодополняющего действия определен-ленных аллелей неаллельных генов, то, очевидно, он будет появляться лишь у тех организмов, которые имеют в генотипе именно такую комбинацию аллелей.
Например, присутствие в генотипе доминантных аллелей обоих неаллельных генов обеспечивает развитие сложного признака, чего не происходит при отсутствии одного из них в доминантном состоянии. В этом случае при скрещивании двух дигетерозиготных организмов, имеющих данный признак, лишь у определенной части потомства (9/16) будет формироваться такой признак, а у остальных (7/16) он не разовьется (рис. 6.17).
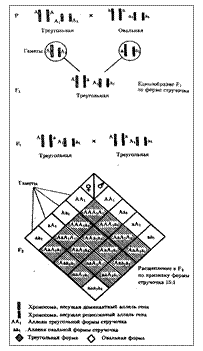
Рис. 6.16. Полимерное наследование формы стручочка у пастушьей сумки
Возможна также ситуация, когда каждый из неаллельных генов в отсутствие доминантного аллеля другого обеспечивает развитие определенного варианта
признака, а вместе они формируют новый его вариант (рис. 6.18). Тогда расщепление в потомстве двух дигетерозигот будет соответствовать расщеплению при независимом наследовании признаков (9:3:3:1).
У человека два гена, детерминирующих отложение в волосах черного и красного пигментов; при определенных сочетаниях их аллелей обеспечивают появление нового признака — особого блеска волос.

Рис. 6.17. Комплементарное взаимодействие генов I
(наследование признака окраски лепестков у душистого горошка)
Наследование признаков при эпистатическом взаимодействии генов.
При эпистазе один из генов (В) выражается фенотипически лишь при отсутствии в генотипе определенного аллеля другого гена (А). В его присутствии действие гена В не проявляется. В строгом смысле слова, этот вид взаимодействия неаллельных генов может быть рассмотрен как вариант взаимодополняющего действия определенных аллелей этих генов, когда один из них способен обеспечить развитие признака, но лишь в присутствии определенного аллеля другого гена. В этой ситуации фенотип организма зависит от конкретного сочетания аллелей неаллельных генов в их генотипах и расщепление по фенотипу в потомстве двух дигетерозигот по этим генам может быть различным.
Рис. 6.18. Комплементарное взаимодействие генов II
(наследование формы гребня у кур):
I — А?bb —розовидная, II — ааВ? — гороховидная, III — А?В? —ореховидная, IV — aabb —листовидная
При доминантном эпистазе, когда доминантный аллель одного гена (А) препятствует проявлению аллелей другого гена (В или b), расщепление в потомстве зависит от их фенотипического значения и может выражаться соотношениями 12:3:1 или 13:3 (рис. 6.19). При рецессивном эпистазе ген, определяющий какой-то признак (В), не проявляется у гомозигот по рецессивному аллелю другого гена (аа). Расщепление в потомстве двух дигетерозигот по таким генам будет соответствовать соотношению 9:3:4 (рис. 6.20). Невозможность формирования признака при рецессивном эпистазе расценивают также как проявление несостоявшегося комплементарного взаимодействия, которое возникает между доминантным аллелем эпистатического гена и аллелями гена, определяющего этот признак.
С этой точки зрения может быть рассмотрен «Бомбейский феномен» у человека, при котором у организмов-носителей доминантного аллеля гена, определяющего группу крови по системе АВ0 (IA или IB), фенотипически эти аллели не проявляются и формируется I группа крови (см. рис. 3.82). Отсутствие фенотипического проявления доминантных аллелей гена I связывают с гомозиготностью некоторых организмов по рецессивному аллелю гена Н (hh), что препятствует формированию антигенов на поверхности эритроцитов. В браке дигетерозигот по генам Н и I (НhIAIB) 1/4 потомства будет иметь фенотипически I группу крови в связи с их гомозиготностью по рецессивному аллелю гена Н — hh.
Рассмотренные выше расщепления по фенотипу в потомстве от скрещивания гетерозиготных родителей или анализирующего скрещивания как при моногенном типе наследования признаков, так и в случае взаимодействия неаллельных генов носят вероятностный характер. Такие расщепления наблюдаются лишь в том случае, если реализуются все возможные встречи разнообразных гамет при оплодотворении и все потомки оказываются жизнеспособными. Выявление близких расщеплений вероятно при анализе большого количества потомков, когда случайные события не способны изменить характер расщепления. Г. Мендель, разработавший приемы гибридологического анализа, впервые применил статистический подход к оценке получаемых результатов. Он анализировал большое число потомков, поэтому расщепления по фенотипу, наблюдаемые им в опытах, оказались близкими к расчетным, которые получаются при учете всех типов гамет, образуемых в мейозе, и их встреч при оплодотворении.
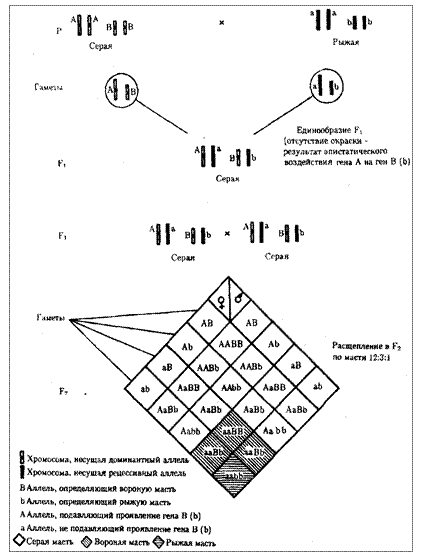
Рис. 6.19. Эпистатическое взаимодействие генов. Доминантный эпистаз (наследование масти у лошадей)
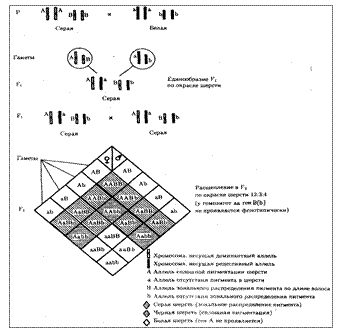
Рис. 6.20. Эпистатическое взаимодействие генов. Рецессивный эпистаз (наследование пигментации шерсти у мышей)
Дата добавления: 2015-06-01; просмотров: 2552;