Структурная организация фотосинтетического аппарата, строение листа как органа фотосинтеза, ультраструктура и онтогенез хлоропластов
Основные принципы действия регуляторных механизмов растительной клетки; компартментация каталитических систем и метаболических фондов протопласта как один из механизмов регуляции клеточного метаболизма.
Клетка – это мельчайшая единица жизни, характеризующаяся определенным типом обмена веществ, самостоятельным энергетическим циклом и способностью к саморегуляции и саморазвитию на основе взаимодействия различных своих компо-нентов и взаимодействия с внешними условиями. В живой клетке нет изолированных процессов. Основой этого взаимодействия служит непрерывный обмен продуктами реакций, образующимися в органоидах определенного вида и поступающими в цитоплазму и другие органоиды.
Сложное строение растительной клетки, дифференцированной на большое количество специализированных органоидов и структур в соответствии с выполняемыми ее функциями, требует и совершенных систем регуляции. Регуляция обеспечивает целостность и гомеостаз организма, т. е. сохранение постоянства параметров внутренней среды, а также создает условия для его развития (эпигенеза).
В ходе эволюции сначала возникли внутриклеточные системы регуляции. К ним относятся регуляция на уровне ферментов, генетическая и мембранная регуляции. Все эти системы регуляции тесно связаны между собой. Принципы действия регуляторных механизмов клетки:
– рецепторно-конформационный принцип;
– принцип компартментации;
– принцип взаимодействия компонентов цитоплазмы.
В основе всех форм внутриклеточной регуляции лежит единый первичный принцип – рецепторно-конформационный принцип. Белковая молекула – фермент, рецептор или регуляторный белок — «узнает» специфический для нее фактор и, взаимодействуя с ним, изменяет свою конфигурацию. В мультикомпонентных комплексах ферментов, генов и мембран конформационные изменения молекул-рецепторов кооперативно передаются на весь комплекс, влияя на его функциональную активность.
Важнейшими в регуляторных механизмах клетки является принцип компар-тментации, обуславливающий высокую степень временной и пространственной ор-ганизованности метаболизма, и взаимодействия компонентов цитоплазмы. Взаимо-действие обеспечивается наличием гиалоплазмы и эндоплазматической сети.
Компартменты – дифференцированные, специализированные участки, или отсеки в протоплазме, различающиеся по степени активности содержащихся в них химических соединений и систем, регулирующих их превращения. Явление компартментации обуславливают клеточные мембраны, которые выполняют функцию расчленения биохимических процессов, разделения их между различными компонентами протоплазмы и пространственного размещения в объеме клетки фондов метаболитов и ферментов.
В растительной клетке имеется три основных компартмента: свободное пространство (СП), цитоплазма и вакуоль. Компартментальны отдельные участки клетки и органелл, отдельные участки мембран, отдельные фонды метаболитов, ферментов и всех других морфологических и химических компонентов протопласта.
Характерная особенность компартментированных метаболитов и ферментов — их подвижность, что достигается включением метаболитов в везикулы и мелкие вакуоли и концентрацией их в трубочках эндоплазматического ретикулума и других трубчатых структурах. Это способствует, например, отводу ассимилятов из хлоропластов без их соприкосновения со стромой цитоплазмы. Эффективность действия механизмов фотофосфорилирования в хлоропластах определяется компартментами, обеспечивающими образование протонного градиента. Компартментальны в данном случае мембранные диски гран хлоропластов. Практически все основные пути метаболизма углеводов прочно ассоциированы с определенными участками плазматических мембран. В клетке существуют компартменты, содержащие системы гликолиза, цикла Кребса и пентозофосфатного шунта. Так все участки цикла Кребса локализованы в матриксе или во внутренней мембране митохондрий. Аналогичные закономерности установлены для процессов биосинтеза и превращения аминокислот, поглощения и выделения минеральных веществ, воды, регуляции действия имеющихся в клетке ферментов и т. п.
С появлением многоклеточных организмов развиваются и совершенствуются межклеточные системы регуляции. Они включают в себя по крайней мере трофическую, гормональную и электрофизиологическую системы. Такие взаимодействия сразу обнаруживаются при культивировании частей и органов растений в изолированном виде. Для поддержания жизни изолированных частей в инкубационную среду необходимо добавлять трофические и гормональные факторы, в норме поступающие из других органов целого растения.
Важным также является взаимная координация процессов, реакций, координация накопления и выделения веществ и энергии клеткой. Так, нуклеиновые кислоты играют роль хранителя и передатчика наследственной информации (ДНК) и посредника ее реализации (РНК). Белки несут функции рабочих механизмов – это ферменты, которые входят в состав клеточных структур и многих активных соединений. Основной механизм регуляции внутриклеточных процессов связан с влиянием на ферменты разнообразных факторов. Это может осуществляться на генетическом уровне, если определяется состав ферментов или количество того или иного фермента в клетке. Второй тип регуляции – влияние на сам фермент, в результате чего активность его может стимулироваться или ингибироваться. Может иметь место также структурный уровень регуляции – влияние на образование и сборку клеточных структур (мембран, хлоропластов, рибосом, митохондрий и т. д.). Конкретными регуляторами внутриклеточных процессов могут быть фитогормоны, которые вырабатываются клетками или окружающими их другими клетками, или продукты реакций по принципу обратной связи (аллостерический эффект). Регуляция может осуществляться также путем транспортировки в клетку продуктов метаболизма и их предшественников, ионов, изменения формы регулируемого фермента (конформаций).
Структурная организация фотосинтетического аппарата, строение листа как органа фотосинтеза, ультраструктура и онтогенез хлоропластов
«Зеленый лист, или, вернее, микроскопическое зеленое зерно хлорофилла является фокусом, точкой в мировом пространстве, в которую с одного конца притекает энергия солнца, а с другого берут начало все проявления жизни на земле. Растение — посредник между небом и землею. Оно истинный Прометей, похитивший огонь с неба» ( К. А. Тимирязев ).
Основой фотосинтетического аппарата служат:
− у высших зелёных растений внутриклеточные органеллы – хлоропласты;
− у большинства водорослей − специализированные внутриклеточные органеллы – хроматофоры;
− у фотосинтезирующих бактерий и синезелёных водорослей – тилакоиды (мембраны их содержат пигмент бактериохлорофилл или бактериовиридин, а также др. компоненты реакций фотосинтеза), погруженные в периферические слои цитоплазмы.
Хлоропласты занимают 20–30% объёма растительной клетки. У водорослей, например хламидомонады, имеется один хлоропласт, в клетке высших растений содержится от 10 до 70 хлоропластов и более.
Хлорофилл и каротиноиды, погруженные в тилакоидные мембраны хлоропластов, собраны в функциональные единицы – фотосистемы, каждая из которых содержит примерно 250 молекул пигментов. Фотосистема – совокупность ССК (светособирающего комплекса), фотохимического реакционного центра и переносчиков электрона.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины, за ними фикоцианины и аллофикоцианины, последовательно передающие энергию на хлорофилл a реакционного центра.
Роль ССК состоит в том, чтобы собирать и передавать энергию квантов света на небольшое количество молекул (молекулы-ловушки) реакционных центров П680 и П700 которые и осуществляют фотохимические реакции.
Органом фотосинтеза у высших растений является лист. Кроме фотосинтеза в жизни растений лист выполняет функции газообмена, транспирации, синтеза ряда органических веществ, в том числе и фитогормонов.
Листья, будучи боковыми органами, как правило, имеют более или менее плоскую форму, что способствует созданию максимальной фотосинтезирующей поверхности.
Основной тканью листа является паренхимный комплекс клеток, несущий хлоропласты – мезофилл (греч. mesos – средний и филл) (рисунок 1). Остальные ткани листа обеспечивают работу мезофилла и поддерживают связь с окружающей средой. Покровная ткань (эпидермис) регулирует газообмен и транспирацию, защищает лист от внешних воздействий. Проводящие ткани осуществляют отток и приток веществ, поддерживают нормальное оводнение фотосинтезирующих клеток. Механические ткани совместно с живыми тургесцентными клетками мезофилла и эпидермиса образуют опорную систему листа.
Мезофилл. Представлен клетками основной паренхимы, расположенными между верхним и нижним эпидермисом (исключая механические ткани и проводящие пучки). Состоит из живых клеток, с тонкими оболочками, округлой или слегка вытянутой формы, реже с небольшими выростами. Благодаря многочисленным хлоропластам мезофилл окрашен в зеленый цвет, так называемая хлоренхима листа. Иногда оболочки клетки образуют складки внутрь полости клетки, что увеличивает рабочую поверхность хлоренхимных клеток.
У типичных дорсовентральных листьев мезофилл неоднороден и диффе-ренцирован на столбчатую (палисадную) и губчатую паренхиму.
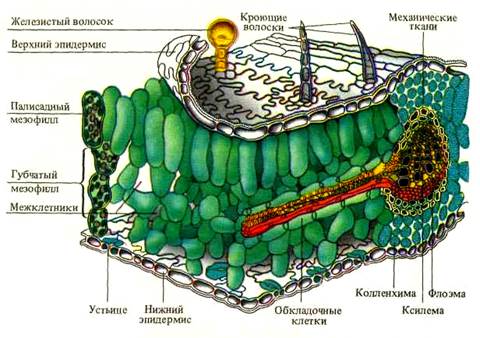
Рисунок 1 – схема строения листа
Клетки столбчатой паренхимы плотно сомкнуты, имеют продолговатую форму и располагаются перпендикулярно к верхней стороне листа. Столбчатый мезофилл может быть однослойный, иногда двуслойный, редко (у светолюбивых растений) многослойный. Основная функция палисадной паренхимы – ассимиляция углекислого газа (фотосинтез). Палисадная ткань располагается в наилучших условиях освещения, непосредственно под верхней эпидермой. Благодаря тому, что ее клетки вытянуты перпендикулярно к поверхности листа, у них имеется возможность регулировать направление и расположение хлоропластов таким образом, чтобы избежать повреждающего действия прямой солнечной радиации на фотосинтезирующий аппарат.
Губчатую паренхиму составляют относительно округлые клетки с большими межклетниками, которые занимают больший объем, чем сами клетки. Рыхлая структура мезофилла обусловливает большую общую площадь поверхности клеток, обращенной к межклетникам. Через крупные межклетники губчатого мезофилла идет газообмен.
Хлоропласты
Форма – овальные тельца длиной 5-10 мкм и диаметром 2-3 мкм. Количество в клетке листа – 15-20, у некоторых водорослей - 1-2 гигантских хлоропласта различной формы. Окраска – зеленая, обусловлена содержанием в хлоропластах хлорофилла.
Хлоропласты – двухмембранные органоиды (рисунок 2). Внутреннее содержимое хлоропластов − строма (матрикс) представляет собой гомогенную среду. Хлоропласты, как и другие органоиды, например, митохондрии, имеют ламеллярное строение, т.е. внутренние структуры органоида образованы инвагинациями внутренней мембраны – ламеллами. В хлоропластах внутренняя мембрана образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран.
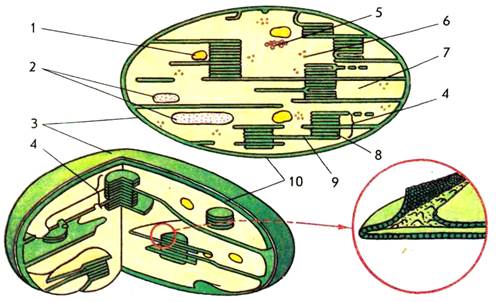
Рисунок 2 – схема строения хлоропласта:
1 – липидная капля; 2 – зерна крахмала; 3 – наружная мембрана; 4 – грана; 5 – ДНК; 6 – рибосома; 7 – строма; 8 – тилакоид граны; 9 – тилакоид стромы; 10 – внутренняя мембрана
Несколько лежащих друг над другом тилакоидов образуют стопку – грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы. При таком строении значительно увеличивается фотоактивная поверхность хлоропластов и обеспечивается максимальное использование световой энергии. В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редокс-цепей и запасания энергии, участвующие в поглощении и использовании энергии света.
Граны часто имеют форму цилиндров размером 2 мкм, в которых пигментно-липидные слои чередуются с белковыми слоями. Морфологически тилакоиды не гомогенны. На внутренней поверхности их мембран имеются специфические структурные образования, названные Парком квантосомами.
Хлоропласты – системы, способные к автономному синтезу белков. В них присутствуют низко- и высокомолекулярная РНК, специфическая кольцевая ДНК и ферменты, активирующие аминокислоты. Хлоропласты обладают собственными рибосомами.
Функции хлоропластов:
1) осуществление фотосинтеза (основная функция);
2) в хлоропластах происходит вся сложная цепь процессов превращений первичных продуктов фотосинтеза (наращивание углеродной цепи, образование и распад полимерных форм углеводов и др.);
Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал. В хлоропластах представлен весь набор биохимических систем, участвующих в синтезе АТФ.
Онтогенез хлоропластов. При нормальном освещении пропластиды пре-вращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид – этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов.
Дата добавления: 2015-05-30; просмотров: 4266;