ВОДНЫЙ ОБМЕН
3.1. Значение воды для растения
Вода является главной составной частью растений. Ее содержание неодинаково в разных органах растения (так, в листьях салата она составляет 95 %, а в сухих семенах - не более 10 % от массы ткани) и зависит от условий внешней среды, вида и возраста растения. Для своего нормального существования растение должно содержать определенное количество воды. Два процесса – поступление и испарение воды – называют водным балансом.
Вода - это среда, в которой протекают процессы обмена веществ. Все реакции гидролиза, окислительно-восстановительные реакции идут с участием воды. Вода служит источником кислорода, выделяемого при фотосинтезе, и водорода, используемого для восстановления углекислого газа. Вода поддерживает конформацию молекул белка, устойчивость структур цитоплазмы и оболочки клеток в упругом состоянии. С изменением тургорного давления связаны некоторые движения частей растений.
Заряды в молекуле воды распределены неравномерно, так как атом кислорода воды оттягивает электроны от атомов водорода. Поэтому молекула воды представляет собой диполь: один полюс молекулы заряжен положительно, а другой отрицательно. Благодаря этому молекулы воды могут ассоциировать друг с другом, ионами и белковыми молекулами. Вода участвует в поглощении и транспорте веществ, так как является хорошим растворителем. Гидратные оболочки, окружающие ионы, ограничивают их взаимодействие.
Вода обладает высокой теплоемкостью - 1кал/град, что позволяет растению воспринимать изменения температуры окружающей среды в смягченном виде. Испарение воды растениями - транспирация служит основным средством терморегуляции у растений. Растения испаряют очень много воды. Большой расход воды связан с тем, что растения обладают значительной листовой поверхностью, необходимой для поглощения углекислого газа, содержание которого в воздухе незначительно (0,032 %).
3.2. Формы почвенной влаги
По степени доступности для растения различают следующие формы почвенной влаги. Гравитационная вода заполняет промежутки между частицами почвы и хорошо доступна растениям. Однако она быстро испаряется и легко стекает в нижние горизонты почвы под влиянием силы тяжести, вследствие чего бывает в почве лишь после дождей. Капиллярная вода заполняет капилляры в почвенных частицах. Эта вода хорошо доступна для растений, она удерживается в капиллярах силами поверхностного натяжения и поэтому не только не стекает вниз, но и поднимается вверх от грунтовых вод. Пленочная вода окружает коллоидные частицы почвы. Вода из периферических слоев гидратационных оболочек может поглощаться корнями. Гигроскопическая вода адсорбируется сухой почвой при помещении ее в атмосферу с 95 %-ной относительной влажностью. Этот тонкий слой молекул воды удерживается с такой силой, что их водный потенциал достигает -1000 бар и она недоступна для растений.
Количество почвенной воды в процентах, при котором растение впадает в устойчивое завядание, называют коэффициентом или влажностью завядания. Завядание растений разных видов может начинаться при одной и той же влажности, но промежуток времени от завядания растения до его гибели (интервал завядания) у растений может быть различным. Так, для растений бобов он составляет несколько суток, а для растений проса - несколько недель. Завядание начинается позже у растений с более отрицательным осмотическим потенциалом и меньшей скоростью транспирации.
«Мертвый запас» влаги в почве - это количество воды полностью недоступной растению. Он зависит от механического состава почвы. Чем больше глинистых частиц в почве, тем больше «мертвый запас» влаги. Количество доступной для растения воды представляет собой разность между полевой влагоемкостью (максимальное количество воды, удерживаемое почвой) и «мертвым запасом».
3.3. Формы воды в растении
Вода в растении состоит из фракций, различающихся по своей подвижности из-за связи с различными соединениями. 85-90 % воды приходится на более подвижную фракцию. В эту фракцию входит резервная вода, заполняющая вакуоли и другие компартменты клетки. Она осмотически связана с сахарами, органическими кислотами, минеральными солями и другими растворенными в ней веществами. Осмотически связанной водой называют воду, образующую периферические слои гидратационных оболочек вокруг ионов и молекул. К подвижной фракции относят и интерстициальную воду, выполняющую транспортную функцию и находящуюся в клеточных стенках, межклетниках и сосудах растения.
Фракция малоподвижной воды составляет 10-15 % всей воды клетки. Это конституционная вода, химически связанная и входящая в состав неорганических соединений, а также гидратационная вода, образующая оболочки вокруг молекул веществ. Воду, гидратирующую мицеллы, называют коллоидносвязанной. Молекулы воды располагаются вокруг мицеллы несколькими слоями. Ближайший к поверхности мицеллы слой воды очень прочно связан. За этим слоем следуют все менее прочно связанные слои, молекулы которых могут обмениваться с молекулами свободной воды. Коллоидносвязанная воданеобходима для нормального функционирования клетки и ее устойчивости при попадании в неблагоприятные условия.Коллоидные мицеллы могут гидратироваться не только путем присоединения молекул воды к гидрофильным группам, расположенным на поверхности - это так называемая мицеллярная гидратация, но и путем внедрения молекул воды внутрь мицеллы и присоединения к имеющимся здесь активным гидрофильным радикалам. Такая гидратация называется пермутоидной.
3.4. Корневая система как орган поглощения воды
Наземные растения, в основном, поглощают воду из почвы. Однако некоторое количество воды может попадать в листья из воздуха. Есть даже растения, для которых атмосфера является главным источником влаги. Это эпифиты, живущие на поверхности других растений, но не являющиеся паразитами. Они обладают воздушными корнями с полыми тонкостенными клетками и впитывают парообразную влагу и воду осадков подобно губке. У некоторых эпифитов дождевая вода собирается листьями и затем всасывается с помощью листовых волосков.
Корневая система является органом поглощения воды из почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной структурой. Подсчитано, что общая поверхность корневой системы может превышать поверхность надземных органов примерно в 150 раз. Рост корня и его ветвление продолжаются в течение всей жизни растения.
Поглощение воды и питательных веществ осуществляется корневыми волосками ризодермы. Ризодерма - это однослойная ткань, покрывающая корень снаружи. У одних видов растений каждая клетка ризодермы формирует корневой волосок, у других она состоит из двух типов клеток: трихобластов, образующих корневые волоски, а атрихобластов, не способных к образованию волосков.
Из ризодермы вода попадает в клетки коры. У травянистых растений кора корня обычно представляет собой несколько слоев живых паренхимных клеток. Между клетками имеются крупные межклетники, обеспечивающие аэрацию корня. Через клетки коры возможны два пути транспорта воды и растворов минеральных солей: по симпласту и апопласту. Более быстрый транспорт воды происходит по апопласту, так как в цитоплазме вода отвлекается на нужды клетки.
Затем вода попадает в клетки эндодермы. Эндодерма - это внутренний слой клеток коры, граничащий с центральным цилиндром. Их клеточные стенки водонепроницаемы из-за отложения суберина и лигнина (пояски Каспари). Поэтому вода и соли проходят через клетки эндодермы по симпласту и транспорт воды в эндодерме замедляется (рис. 3.1). Это необходимо, так как диаметр стели (центрального цилиндра), куда попадает вода из эндодермы, меньше всасывающей поверхности корня.
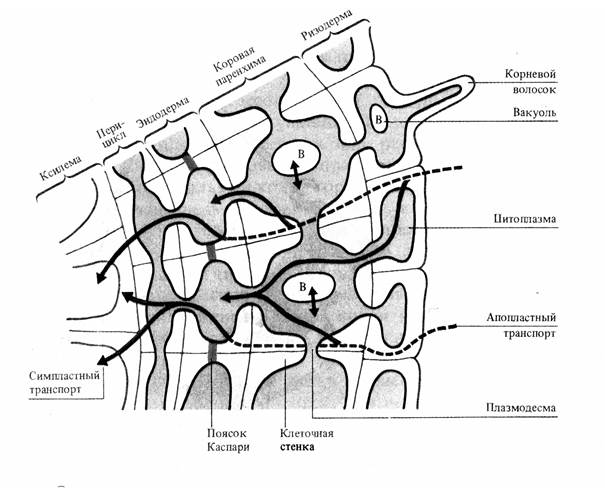
Рис. 3.1. Схема путей радиального транспорта воды и ионов через корень до сосудов ксилемы (по J. Moorby, 1981 – цит. по В. В. Полевому).
Центральный цилиндр корня содержит перицикл и две системы проводящих элементов: ксилему и флоэму. Клетки перицикла представляют собой одно- или многослойную обкладку проводящих сосудов. Его клетки регулируют транспорт веществ как из наружных слоев в ксилему, так и из флоэмы в кору. Кроме того, клетки перицикла выполняют функцию образовательной ткани, способной продуцировать боковые корни. Паренхимные клетки перицикла активно транспортируют ионы в проводящие элементы ксилемы. Контакт осуществляется через поры во вторичных клеточных стенках сосудов и клеток. Между ними нет плазмодесм. Затем вода и растворенные вещества диффундируют в полость сосуда через первичную клеточную стенку. Для некоторых паренхимных клеток сосудистого пучка характерны выросты - лабиринты стенок, выстланные плазмалеммой, что значительно увеличивает ее площадь. Эти клетки активно участвуют в транспорте веществ в сосуды и обратно и называются передаточными или переходными. Они могут граничить одновременно с сосудами ксилемы и ситовидными трубками флоэмы. По сосудам флоэмы транспортируются органические вещества из надземной части растения в корни.
Вода пассивно диффундирует в сосуды ксилемы благодаря осмотическому механизму. Осмотически активными веществами в сосудах являются минеральные ионы и метаболиты, выделяемые насосами плазмалеммы паренхимных клеток, окружающих сосуды. Сосущая сила сосудов выше, чем у окружающих клеток из-за повышающейся концентрации ксилемного сока и отсутствия значительного противодавления со стороны малоэластичных клеточных стенок. В результате поступления воды в сосудах ксилемы развивается гидростатическое давление, получившее название корневого давления. Оно участвует в поднятии ксилемного раствора по сосудам ксилемы из корня в надземную часть растения. Поднятие воды по растению вследствие развивающегося корневого давления называют нижним концевым двигателем.
Примером работы нижнего концевого двигателя служат плач растений. Весной у кустарников и деревьев с еще нераспустившимися листьями можно наблюдать интенсивный ксилемный ток снизу вверх через надрезы ствола и веток. У травянистых растений при отрезании стебля из пенька выделяется ксилемный сок, называемый пасокой.
Поступление воды через корневую систему сокращается с понижением температуры. Это происходит по следующим причинам: 1) повышается вязкость воды и поэтому снижается ее подвижность, 2) уменьшается проницаемость протоплазмы для воды, 3) тормозится рост корней, 4) уменьшается скорость метаболических процессов. Поступление воды снижается при ухудшении аэрации почвы. Это можно наблюдать, когда после сильного дождя почва залита водой, но при ярком солнце из-за сильного испарения растения завядают. Большое значение имеет концентрация почвенного раствора. Вода поступает в корень только тогда, когда водный потенциал корня меньше водного потенциала почвы. Если почвенный раствор имеет более отрицательный потенциал, вода не будет поступать в корень, а выходить из него.
3.5. Передвижение воды по сосудистой системе
Восходящий поток воды в растении идет по сосудам ксилемы, лишенным цитоплазмы. Помимо работы нижнего концевого двигателя и присасывающего действия транспирации (верхний концевой двигатель) в передвижении воды по капиллярным сосудам ксилемы участвуют силы сцепления (когезии) молекул воды друг с другом и силы прилипания (адгезии) воды к стенкам сосудов. Обе силы препятствуют также образованию пузырьков воздуха, способных закупорить сосуд. Скорость передвижения воды по ксилеме равна 12-14 м/ч.
Большая часть воды, попавшей в листья, испаряется в атмосферу, а меньшая часть (около 0,2 %) используется в метаболизме клеток, на поддержание тургора и в транспорте органических соединений по сосудам флоэмы. Вода из клеток листа и непосредственно из сосудов ксилемы поступает во флоэмные окончания по осмотическому градиенту, возникающему вследствие накопления в клетках флоэмы сахаров и других органических соединений, которые образуются в клетках листьев и переносятся в клетки флоэмы в результате активной работы транспортных насосов. Нисходящий флоэмный ток доставляет органические соединения тканям корня, где они используются в метаболизме. В корне окончания проводящих пучков флоэмы, как и в листе, располагаются вблизи элементов ксилемы и вода по осмотическому градиенту поступает в ксилему и движется вверх с восходящим током. Таким образом происходит обмен воды в проводящей системе корня и листьев.
3.6. Транспирация
Транспирация - это испарение воды растением. Основным органом транспирации является лист. Вода испаряется с поверхности листьев через клеточные стенки эпидермальных клеток и покровные слои (кутикулярная транспирация) и через устьица (устьичная транспирация). В результате потери воды в ходе транспирации в клетках листьев возрастает сосущая сила. Это приводит к усилению поглощения клетками листа воды из сосудов ксилемы и передвижению воды по ксилеме из корней в листья. Таким образом, верхний концевой двигатель, участвующий в транспорте воды вверх по растению, обусловлен транспирацией листьев. Верхний концевой двигатель может работать при полном отключении нижнего концевого двигателя, причем для его работы используется не только метаболическая энергия как в корне, но и энергия внешней среды - температура и движение воздуха.
Транспирация спасает растение от перегрева. Температура сильно транспирирующего листа может примерно на 7 Со быть ниже температуры нетранспирирующего завядшего листа. Кроме того, транспирация участвует в создании непрерывного тока воды с растворенными минеральными и органическими соединениями из корневой системы к надземным органам растения.
Транспирацию обычно выражают в следующих единицах. Интенсивность транспирации - это количество воды, испаряемой растением в г за единицу времени в часах единицей поверхности в дм2. Эта величина колеблется от 0,15 до 1,5. Транспирационный коэффициент - это количество воды в г, испаряемой растением при накоплении им 1 г сухого вещества. Продуктивность транспирации - это величина, обратная транспирационному коэффициенту и равна количеству сухого вещества в г, накопленного растением за период, когда оно испаряет 1 кг воды. Относительная транспирация - это отношение воды, испаряемой листом, к воде, испаряемой со свободной водной поверхности той же площади за один и тот же период времени. Экономность транспирации - это количество испаряемой воды в мг на 1 кг воды, содержащейся в растении.
Кутикулярная транспирация. Снаружи листья имеют однослойный эпидермис, внешние стенки клеток которого покрыты кутикулой и воском, образующие эффективный барьер на пути движения воды. На поверхности листьев часто развиты волоски, которые также влияют на водный режим листа, так как снижают скорость движения воздуха над его поверхностью и рассеивают свет и тем самым уменьшают потери воды за счет транспирации. Интенсивность кутикулярной транспирации варьирует у разных видов растений. У молодых листьев с тонкой кутикулой она может составлять около половины всей транспирации. У зрелых листьев с более мощной кутикулой кутикулярная транспирация равна 1/10 общей транспирации. В стареющих листьях из-за повреждения кутикулы она может возрастать. Таким образом, кутикулярная транспирация регулируется главным образом толщиной и целостностью кутикулы и других защитных покровных слоев на поверхности листьев.
Устьичная транспирация. Устьица представляют собой щель в подъустьичную полость, окаймленную двумя замыкающими клетками серповидной формы. Устьица играют важную роль в газообмене между листом и атмосферой, так как являются основным путем для водяного пара, углекислого газа и кислорода. Устьица находятся на обеих сторонах листа. Есть виды растений, у которых устьица располагаются только на нижней стороне листа. В среднем число устьиц колеблется от 50 до 500 на 1 мм2. Транспирация через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом И. Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий мала по отношению к площади всего листа (0,5-2 %), испарение воды через устьица идет очень интенсивно.
Транспирация слагается из двух процессов: 1) передвижения воды в листе из сосудов ксилемы по симпласту и, преимущественно, по клеточным стенкам, так как в стенках транспорт воды встречает меньшее сопротивление, 2) испарения воды из клеточных стенок в межклетники и подъустьичные полости с последующей диффузией в окружающую атмосферу через устьичные щели. Чем меньше относительная влажность атмосферного воздуха, тем ниже его водный потенциал. Если водный потенциал воздуха меньше водного потенциала подъустьичных полостей, то молекулы воды испаряются наружу.
Основным фактором, влияющим на открывание и закрывание устьиц, является содержание воды в листе, в том числе и в замыкающих клетках устьиц. Клеточные стенки замыкающих клеток имеют неодинаковую толщину. Внутренняя часть стенки, примыкающая к устьичной щели, более толстая, а внешняя – более тонкая. По мере того как замыкающая клетка осмотически поглощает воду, более тонкая и эластичная часть ее клеточной стенки растягивается и оттягивает внутреннюю часть стенки. Замыкающие клетки принимают полукруглую форму и устьица раскрываются. При недостатке воды замыкающие клетки выпрямляются и устьичная щель закрывается (рис. 3.2). Кроме того, по мере увеличения водного дефицита в тканях растения повышается концентрация ингибитора роста абсцизовой кислоты. Она подавляет деятельность Н+-насосов в плазмалемме замыкающих клеток, вследствие чего снижается их тургор и устьица закрываются. Абсцизовая кислота также ингибирует синтез фермента a-амилазы, что приводит к снижению гидролиза крахмала. По сравнению с низкомолекулярными углеводами крахмал не является осмотически активным веществом, поэтому сосущая сила замыкающих клеток уменьшается и устьица закрываются.
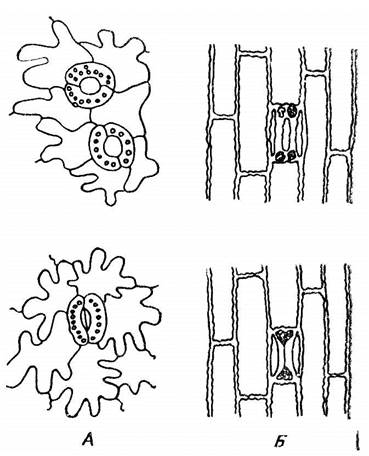
Рис. 3.2. Устьица в открытом (вверху) и закрытом (внизу) состоянии.
А – двудольного растения, Б – злака (по С. И. Лебедеву).
В отличие от других клеток эпидермиса замыкающие клетки устьиц содержат хлоропласты. Синтез углеводов в процессе фотосинтеза в замыкающих клетках увеличивает их сосущую силу и вызывает поглощение воды, способствуя этим открыванию устьиц.
Состояние устьиц зависит от углекислого газа. Если концентрация СО2 в подъустьичной полости падает ниже 0,03 %, тургор замыкающих клеток увеличивается и устьица открываются. Повышение концентрации СО2 в воздухе вызывает закрытие устьиц. Это происходит в межклетниках листа ночью, когда в результате отсутствия фотосинтеза и продолжающегося дыхания уровень углекислого газа в тканях повышается. Такое влияние углекислого газа объясняет, почему ночью устьица закрыты и открываются с восходом солнца. Сдвиг рН в щелочную сторону вследствие уменьшения концентрации СО2 увеличивает активность ферментов, участвующих в распаде крахмала, тогда как при кислом рН при повышении содержания СО2 в межклетниках повышается активность ферментов, катализирующих синтез крахмала.
На свету замыкающие клетки устьиц содержат значительно больше калия, чем в темноте. При открывании устьиц содержание калия в замыкающих клетках увеличивается в 4 раза при одновременном снижении его содержания в сопутствующих клетках. Установлено повышение содержания АТФ в замыкающих клетках устьиц в процессе их открывания. АТФ, образованная в процессе фотосинтетического фосфорилирования в замыкающих клетках, используется для усиления поступления калия. Усиленное поступление ионов калия повышает сосущую силу замыкающих клеток. В темноте ионы калия выделяются из замыкающих клеток и устьица закрываются.
Периодичность суточного хода транспирации наблюдается у многих растений, но у разных видов растений устьица функционируют неодинаково. У деревьев, теневыносливых растений, многих злаков и других гидростабильных видов с совершенной регуляцией устьичной транспирации испарение воды начинается на рассвете, достигает максимума в утренние часы. В полдень транспирация снижается и вновь увеличивается в предвечерние часы при снижении температуры воздуха. Такой ход транспирации приводит к незначительным суточным изменениям осмотического давления и содержания воды в листьях. У видов растений, способных переносить резкие изменения содержания воды в клетках в течение дня, то есть у гидролабильных видов, наблюдается одновершинный суточный ход транспирации с максимумом в полуденные часы. В обоих случаях ночью транспирация минимальна или полностью прекращается.
3.7. Особенности водного обмена у растений разных экологических групп
Растения, обитающие в воде - гидратофиты или гидрофиты, погружены в воду полностью или частично. Они регулируют постоянство состава внутренней среды с помощью механизмов защиты от избыточного поступления воды. У монадных форм зеленых водорослей, заселяющих, в основном, пресные воды, клеточные стенки замкнуты не полностью из-за наличия выростов цитоплазмы - жгутиков, с помощью которых они передвигаются. У всех монадных форм имеются пульсирующие вакуоли, посредством которых из клеток удаляются избыток воды и отходы жизнедеятельности. У гидрофитов с замкнутой клеточной стенкой ее противодавления достаточно для предотвращения поступления излишков воды в клетку. Первичными гидрофитами являются водоросли. Водные цветковые растения - это вторичные гидрофиты, происходящие от наземных форм.
По способности приспосабливать водный обмен к колебаниям водоснабжения различают две группы наземных растений: пойкилогидрические и гомойгидрические.
Пойкилогидрические организмы (бактерии, синезеленые водоросли, низшие зеленые водоросли, грибы, лишайники и другие) приспособились переносить значительный недостаток воды без потери жизнеспособности. При этом у них снижается интенсивность обмена веществ, клетки равномерно сжимаются. Протопласт их клеток при сильном обезвоживании переходит в состояние геля. Увеличение количества воды в среде приводит к возобновлению активного метаболизма в клетках. По характеру изменения таких показателей водного режима, как интенсивность транспирации, осмотическое давление, содержание воды в течение суток они относятся к гидролабильным растениям, так как у них значительно изменяются содержание воды и испарение.
Гомойгидрические растения (наземные папоротникообразные, голосеменные, цветковые) составляют большинство обитателей суши. Они обладают механизмами регуляции устьичной транспирации, а также корневой системой, обеспечивающей доставку воды. Поэтому даже при значительных изменениях влажности среды у этих растений не наблюдается резких колебаний содержания воды в клетках, в которых, как правило, развита вакуолярная система. Их клетки не способны к обратимому высыханию. У этих растений гидростабильный тип водного режима. Стабилизации водного режима у многих видов растений способствуют запасы воды в корнях, стеблях и запасающих органах. Гомойгидрические растения делятся на три экологические группы:
1. Гигрофиты (тонколистные папоротники, некоторые фиалки и другие), произрастающие в условиях повышенной влажности и недостаточной освещенности. Теневыносливые гигрофиты, с почти всегда открытыми устьицами, имеют гидатоды, через которые выделяют избыток воды в капельножидком состоянии. Гигрофиты плохо переносят почвенную и воздушную засуху.
2. Мезофиты (лиственные деревья, лесные и луговые травы, большинство культурных растений) обитают в среде со средним уровнем обеспеченности водой и не имеют ясно выраженных приспособлений к избытку или недостатку воды.
3. Ксерофиты живут в местах с жарким и сухим климатом и приспособлены к перенесению атмосферной и почвенной засухи. Ксерофиты делят на следующие группы:
1. Растения, избегающие засухи (эфемеры). Эти растения обладают коротким вегетационным периодом, приурочивая весь жизненный цикл к периоду дождей и засуху переносят в форме семян.
2. Растения, запасающие влагу (ложные ксерофиты). К этой группе растений относятся суккуленты (кактусы и растения семейства толстянковых). Эти растения живут в районах, где засушливые периоды сменяются периодами дождей. Они имеют толстые и мясистые стебли. Листья часто редуцированы, вся поверхность растений покрыта толстым слоем кутикулы, что существенно снижает их транспирацию. Суккуленты обладают неглубокой, но широко распространяющейся корневой системой. Клетки корня характеризуются сравнительно низкой концентрацией клеточного сока. Вода, запасаемая в мясистых органах, тратится очень медленно.
Суккуленты обладают своеобразным обменом веществ. У них днем устьица закрыты, а ночью они открываются, что обеспечивает снижение расходования воды в процессе транспирации. Углекислый газ поступает через устьица ночью и усваивается с образованием органических кислот. В дневные часы углекислый газ вновь освобождается и используется в процессе фотосинтеза. Поэтому эти растения фотосинтезируют при закрытых днем устьицах. Растения этой группы не устойчивы к длительному водному стрессу.
3. Гемиксерофиты или полуксерофиты - это растения, у которых сильно развиты приспособления к добыче воды. У них глубоко идущая, сильно разветвленная корневая система. Клетки корня обладают высокой концентрацией клеточного сока и очень отрицательным водным потенциалом. Растения этой группы обладают хорошо развитой проводящей системой. Листья у них тонкие, с очень густой сетью жилок, что сокращает путь передвижения воды к клеткам листа. Даже в очень жаркие дни они держат устьица открытыми. Благодаря высокой интенсивности транспирации температура листьев значительно понижается, что позволяет осуществлять фотосинтез при высокой температуре воздуха. Листья некоторых растений покрыты волосками, которые создают экран, дополнительно защищающий листья от перегрева.
4. Эуксерофиты или настоящие ксерофиты - это растения, обладающие способностью резко сокращать транспирацию в условиях недостатка воды. Они имеют приспособления к сокращению потерь воды: подземные органы, а иногда и стебли покрыты толстым слоем пробки, листья покрыты толстым слоем кутикулы, многие имеют волоски, устьица расположены в углублениях, устьичные щели закупорены восковыми и смолистыми пробочками, листья свернуты в трубочку, где создается свой микроклимат и уменьшается контакт устьичных щелей с атмосферой. Для растений этой группы характерна способность переносить обезвоживание и состояние длительного завядания. Особенно хорошо переносят потерю воды растения с жесткими листьями - склерофиты, которые и в состоянии тургора имеют сравнительно мало воды. Эти растения характеризуются большим развитием механических тканей.
Дата добавления: 2015-05-26; просмотров: 1389;