Строение и функции биологических мембран
Биологические мембраны- общее название функционально активных поверхностных структур, ограничивающих клетки (клеточные, или плазматические мембраны) и внутриклеточные органеллы (мембраны митохондрий, ядер, лизосом, эндоплазматического ретикулума и др.). Они содержат в своем составе липиды, белки,гетерогенные молекулы (гликопротеины,гликолипиды)и в зависимости от выполняемой функции многочисленные минорные компоненты: коферменты, нуклеиновые кислоты, антиоксиданты, каротиноиды, неорганические ионы и т. п.
Согласованное функционирование мембранных систем – рецепторов, ферментов, транспортных механизмов - помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
К основным функциям биологических мембран можно отнести:
· отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
· контроль и регулирование транспорта огромного многообразия веществ через мембраны;
· участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
· преобразование энергии пищевых органических веществ в энергию химических связей молекул АТФ.
Молекулярная организация плазматической (клеточной) мембраны у всех клеток примерно одинакова: она состоит из двух слоев липидных молекул с множеством включенных в нее специфических белков. Одни мембранные белки обладают ферментативной активностью, тогда как другие связывают питательные вещества из окружающей среды и обеспечивают их перенос в клетку через мембраны. Мембранные белки различают по характеру связи с мембранными структурами. Одни белки, называемые внешними или периферическими, непрочно связаны с поверхностью мембраны, другие, называемые внутренними или интегральными, погружены внутрь мембраны. Периферические белки легко экстрагируются, тогда как интегральные белки могут быть выделены только при помощи детергенов или органических растворителей. На рис. 4 представлена структура плазматической мембраны.
Внешние, или плазматические, мембраны многих клеток, а также мембраны внутриклеточных органелл, например, митохондрий, хлоропластов удалось выделить в свободном виде и изучить их молекулярный состав. Во всех мембранах имеются полярные липиды в количестве, составляющем в зависимости от типа мембран от 20 до 80% ее массы, остальное приходится главным образом на долю белков. Так, в плазматических мембранах животных клеток количество белков и липидов, как правило, примерно одинаково; во внутренней митохондриальной мембране содержится около 80% белков и только 20% липидов, а в миелиновых мембранах клеток мозга наоборот, около 80% липидов и только 20% белков.
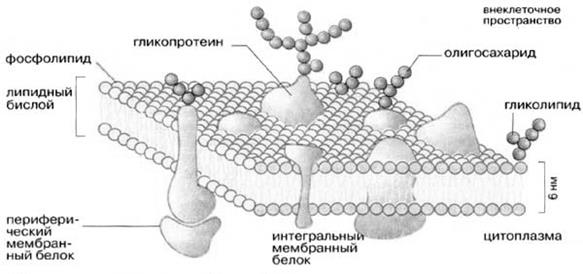
Рис. 4. Структура плазматической мембраны
Липидная часть мембран представляет собой смесь различного рода полярных липидов. Полярные липиды, к числу которых относятся фосфоглицеролипиды, сфинголипиды, гликолипиды не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях.
Все полярные липиды в мембранах постоянно обновляются в процессе метаболизма, при нормальных условиях в клетке устанавливается динамическое стационарное состояние, при котором скорость синтеза липидов равна скорости их распада.
В мембранах животных клеток присутствуют в основном фосфоглицеролипиды и в меньшей степени сфинголипиды; триацилглицеролы обнаруживаются лишь в следовых количествах. Некоторые мембраны животных клеток, в особенности наружная плазматическая мембрана, содержит значительные количества холестерола и его эфиров (рис.5).
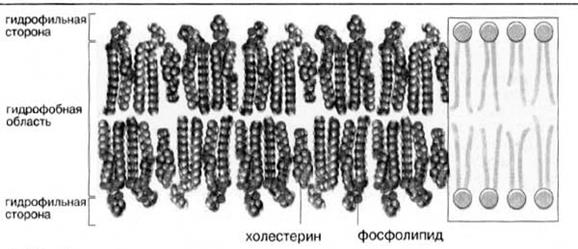
Рис.5. Мембранные липиды
В настоящее время общепринятой моделью строения мембран является жидкостно-мозаичная, предложенная в 1972 году С. Синджером и Дж. Николсоном.
Согласно ей белки можно уподобить айсбергам, плавающим в липидном море. Как уже указывалось выше, существуют 2 типа мембранных белков: интегральные и периферические. Интегральные белки пронизывают мембрану насквозь, они являются амфипатическими молекулами. Периферические белки не пронизывают мембрану и связаны с ней менее прочно. Основной непрерывной частью мембраны, то есть ее матриксом, служит полярный липидный бислой. При обычной для клетки температуре матрикс находится в жидком состоянии, что обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами в гидрофобных хвостах полярных липидов.
Жидкостно-мозаичная модель предполагает также, что на поверхности расположенных в мембране интегральных белков имеются R-группы аминокислотных остатков (в основном гидрофобные группы, за счет которых белки как бы «растворяются» в центральной гидрофобной части бислоя). В то же время, на поверхности периферических, или внешних белков, имеются в основном гидрофильные R-группы, которые притягиваются к гидрофильным заряженным полярным головкам липидов за счет электростатических сил. Интегральные белки, а к ним относятся ферменты и транспортные белки, обладают активностью только в том случае, если находятся внутри гидрофобной части бислоя, где они приобретают необходимую для проявления активности пространственную конфигурацию (рис.6). Следует еще раз подчеркнуть, что ни между молекулами в бислое, ни между белками и липидами бислоя не образуется ковалентных связей.

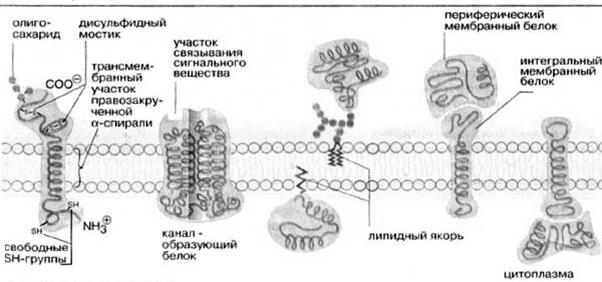
Рис.6. Мембранные белки
Мембранные белки могут свободно перемещаться в латериальной плоскости. Периферические белки буквально плавают на поверхности бислойного «моря», а интегральные белки, подобно айсбергам, почти полностью погружены в углеводородный слой.
В большинстве своем мембраны ассиметричны, то есть имеют неравноценные стороны. Эта ассиметричность проявляется в следующем:
· во-первых, в том, что внутренняя и внешняя стороны плазматических мембран бактериальных и животных клеток различаются по составу полярных липидов. Так, например, внутренний липидный слой мембран эритроцитов человека содержит в основном фосфатидилэтаноламин и фосфатидилсерин, а внешний – фосфатидилхолин и сфингомиелин.
· во-вторых, некоторые транспортные системы в мембранах действуют только в одном направлении. Например, в мембранах эритроцитов имеется транспортная система («насос»), перекачивающая ионы Nа+ из клетки в окружающую среду, а ионы К+ - внутрь клетки за счет энергии, освобождающейся при гидролизе АТФ.
· в-третьих, на внешней поверхности плазматических мембран содержится очень большое число олигосахаридных группировок, представляющих собой головки гликолипидов и олигосахаридные боковые цепи гликопротеинов, тогда как на внутренней поверхности плазматической мембраны олигосахаридных группировок практически нет.
Ассиметричность биологических мембрам сохраняется за счет того, что перенос индивидуальных молекул фосфолипидов с одной стороны липидного бислоя на другую очень затруднен по энергетическим соображениям. Полярная молекула липида способна свободно перемещаться на своей стороне бислоя, но ограничена в возможности перескочить на другую сторону.
Подвижность липидов зависит от относительного содержания и типа присутствующих ненасыщенных жирных кислот. Углеводородная природа жирнокислотных цепей сообщает мембране свойства текучести, подвижности. В присутствии цис-ненасыщенных жирных кислот силы сцепления между цепями слабее, чем в случае одних насыщенных жирных кислот, и липиды сохраняют высокую подвижность и при низкой температуре.
На внешней стороне мембран имеются специфические распознающие участки, функция которых состоит в распознавании определенных молекулярных сигналов. Например, именно посредством мембраны некоторые бактерии воспринимают незначительные изменения концентрации питательного вещества, что стимулирует их движение к источнику пищи; это явление носит название хемотаксиса.
Мембраны различных клеток и внутриклеточных органелл обладают определенной специфичностью, обусловленной их строением, химическим составом и функциями. Выделяют следующие основные группы мембран у эукариотических организмов:
· плазматическая мембрана (наружная клеточная мембрана, плазмалемма),
· ядерная мембрана,
· эндоплазматический ретикулум,
· мембраны аппарата Гольджи, митохондрий, хлорпластов, миелиновых оболочек,
· возбудимые мембраны.
У прокариотических организмов помимо плазматической мембраны существуют внутрицитоплазматические мембранные образования, у гетеротрофных прокариот они называются мезосомами. Последние образуются впячиванием внуть наружной клеточной мембраны и в некоторых случаях сохраняют с ней связь.
Мембрана эритроцитовсостоит из белков (50%), липидов (40%) и углеводов (10%). Основная часть углеводов (93%) связана с белками, остальная – с липидами. В мембране липиды расположены асимметрично в отличие от симметричного расположения в мицеллах. Например, кефалин находится преимущественно во внутреннем слое липидов. Такая асимметрия поддерживается, по-видимому, за счет поперечного перемещения фосфолипидов в мембране, осуществляемого с помощью мембранных белков и за счет энергии метаболизма. Во внутреннем слое эритроцитарной мембраны находятся в основном сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, в наружном слое – фосфатидилхолин. Мембрана эритроцитов содержит интегральный гликопротеин гликофорин, состоящий из 131 аминокислотного остатка и пронизывающий мембрану, и так называемый белок полосы 3, состоящий из 900 аминокислотных остатков. Углеводные компоненты гликофорина выполняют рецепторную функцию для вирусов гриппа, фитогемагглютининов, ряда гормонов. В эритроцитарной мембране обнаружен и другой интегральный белок, содержащий мало углеводов и пронизывающий мембрану. Его называют туннельным белком (компонент а), так как предполагают, что он образует канал для анионов. Периферическим белком, связанным с внутренней стороной эритроцитарной мембраны, является спектрин.
Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около 80%, половина из них – фосфолипидов). Белки этих мембран важны для фиксации лежащих друг над другом мембранных солев.
Мембраны хлоропластов. Хлоропласты покрыты двухслойной мембраной. Наружная мембрана имеет некоторое сходство с таковой у митохондрий. Помимо этой поверхностной мембраны в хлоропластах имеется внутренняя мембранная система – ламеллы. Ламеллы образуют или уплощенные пузырьки – тилакоиды, которые, располагаясь друг над другом, собираются в пачки (граны) или формируют мембранную систему стромы (ламеллы стромы). Ламеллы гран и стромы наружной стороне мембраны тилакоидов сосредоточены гидрофильные группировки, галакто- и сульфолипидов. Фитольная часть молекулы хлорофилла погружена в глобулу и находится в контакте в гидрофобными группами белков и липидов. Порфириновые ядра хлорофилла в основном локализованы между соприкасающимися мембранами тилакоидов гран.
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий. В ней локализованы ферменты дыхательной цепи, активного транспорта; ферменты, участвующие в образовании компонентов мембраны. Преобладающим компонентом бактериальных мембран являются белки: соотношение белок/липид (по массе) равно 3:1. Наружная мембрана грамотрицательных бактерий по сравнению с цитоплазматической содержит меньшее количество различных фосфолипидов и белков. Обе мембраны различаются по липидному составу. Во внешней мембране находятся белки, образующие поры для проникновения многих низкомолекулярных веществ. Характерным компонентом наружной мембраны является также специфический липополисахарид. Ряд белков наружной мембраны служит рецепторами для фагов.
Мембрана вирусов. Среди вирусов мембранные структуры характерны для содержащих нуклеокапсид, который состоит из белка и нуклеиновой кислоты. Это «ядро» вирусов окружено мембраной (оболочка). Она также состоит из двойного слоя липидов с включенными в него гликопротеинами, расположенными в основном на поверхности мембраны. У ряда вирусов (микровирусы) в мембраны входит 70-80% всех белков, остальные белки содержатся в нуклеокапсиде.
Таким образом, мембраны клеток представляют собой очень сложные структуры; составляющие их молекулярные комплексы образуют упорядоченную двумерную мозаику, что придает поверхности мембран биологическую специфичность.
Дата добавления: 2015-05-16; просмотров: 4763;