Наявність світла та
сприятлива температура
СО2 + Н2О + мінеральні = фітомасі + кисень +
речовини + транспірована вода
Фотосинтез суттєво залежить від багатьох ландшафтно-екологічних факторів. З них найбільше значення мають: світло, температура, вода, поживні речовини в грунті. З інтенсивністю світла фотосинтез пов’язаний лінійною залежністю, але до деякого оптимального рівня. Цей рівень називається світловим насиченням і в середньому становить 1 ккал/м2 хв. При його досягненні інтенсивність фотосинтезу може дещо знизитись або лишається сталою. Крім інтенсивності світла, фотосинтез залежить і від тривалості освітлення – чим вона вища, тим більше продукується фітомасі.
Температура в умовах недостатньої освітленості, коли світло є лімітуючим фотосинтез фактором, мало впливає на нього. Проте при помірній освітленості інтенсивність фотосинтезу зростає в 2-5 раз при підвищенні температури на кожні 100С. Залежність між інтенсивністю фотосинтезу та температурою списується кривою дзвоноподібної форми з відносно вузьким діапазоном оптимальних температур. Температурний оптимум у різних видів рослин дуже різний і становить від 160С (для багатьох видів помірного поясу) до 380С (тропічні рослини). Зниження фотосинтезу при високих температурах пояснюється збільшенням витрат енергії на дихання.
Вода як фактор продуктивності виявляє різнобічну, але переважно посередню роль у фотосинтезі. Із зниженням вмісту вологи в грунті рослини зменшують її витрату на транспірацію і тому закривають продихи. Крізь них, крім вивільнення води, рослини поглинають з атмосфери СО2, необхідний для фотосинтезу, тому при закритті продихів цей процес уповільнюється, а при зниженні вологості грунту нижче точки в’янення поступово припиняється. Різні види рослин неоднаково реагують на водний дефіцит, та все ж є тісна залежність між біопродуктивністю та сумарним випаровуванням. Теоретично її обґрунтував Г.Пенман у 1956 р., а емпіричну залежність установив М.Розенцвейг (1968):
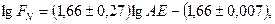
де FN – чиста первинна продукція надземних частин, г/м2;
AE – фактичне річне сумарне випаровування, мм.
Поживні речовини грунту. Для фотосинтезу, крім мікроелементів, необхідні й мікроелементи, особливо Mn, Fe, Cl, Zn, V. Наявність їх у грунті в легкодоступній формі і не в токсичних кількостях сприяє фотосинтезу. Проте в багатьох геосистемах при добрих гідро кліматичних умовах для фотосинтезу його інтенсивність лімітується недостатньою кількістю поживних елементів у грунті, особливо тих, що знаходяться в мінімумі. При внесенні екологічно обґрунтованих норм мінеральних добрив фактор, що обмежує фотосинтез, знімається.
Утворена фітомасі (чиста первинна продукція) далі розподіляється між елементами геосистеми за схемою, аналогічною до потоків енергії по трофічній сітці (див. рис. 7) [1]. Згідно з Р.Уіттекером (1980), частка рослинної продукції, що споживається первинними консументами (травоїдними тваринами), становить в агро екосистемах 1%, у тундрових та пустинних 2-3, лісових 4-7, степових10-15%. І хоч у деяких геосистемах (наприклад, пасовищних) реальний відсоток первинної продукції, споживаної тваринами, може бути набагато більшим, засвоюється вона лише частково (1-15%), а більша її частина виводиться тваринами з екскрементами та при диханні. Тому з переходом на кожний наступний трофічний рівень зоомаси зменшується на 2-3 порядки (див. рис. 11,б) [1].
Таблиця 1.(6 [1]). Середня продуктивність геосистем (за Р.Уіттекером, 1980)
| Тип геосистем | Чиста первинна продукція, т/км2 за рік | Біомаса, т/км2 | Маса під- стилки, т/км2 | Спожи вання тв арина- ми т/км2 за рік | Продук- ція тварин, т/км2 за рік | Біомаса тварин, т/км2 |
| Тундрові Бореальнолісові Широколистолісові Степові Пустинні та напівпустинні Тропічних дощових лісів Болотяні Агрогеосис- теми Усі континенти | 0,38 3,20 6,0 8,9 0,40 15,0 16,0 0,65 6,10 | 0,44 4,75 15,70 6,70 0,45 20,0 10,0 0,40 6,75 |
Продуктивність геосистем та розподіл продукції між окремими ланками трофічної структури досить суттєво змінюються залежно від ландшафтно-екологічних умов (табл.. 6) [1].
Важливою складовою продуційного процесу в геосистемі є продукування мікробної маси. Внаслідок короткої тривалості життя перважної більшості популяцій мікроорганізмів (15-20 днів і менше) в помірному поясі за рік змінюється 6-7 їх поколінь, а в тропічному – більше 20. Окремі популяції бактерій дають за рік 30-40 генерацій і більше (Кожевін, 1973). Така велика швидкість відновлення біомаси призводить до значної величини її річної продукції. Так, за даними М.Віткам па (1974), у лісовому опаді добова продукція грибного міцелію становить 1 г/м2, що приблизно відповідає продуктивності зернових культур. Майже такі самі оцінки продуктивності наводить Т.В.Арістовська (1980) і для мікроорганізмів грунту – від 0,15 до 3,9 мг/г за добу. За даними В.А.Ковди та І.В.Янушевської (1973), щорічна продукція мікроорганізмів становить 200-500 ц/га, що набагато перевищує продуктивність вищих рослин.
Характерна особливість продуційного процесу мікроорганізмів полягає в чітко вираженій його пульсації – протягом року періоди інтенсивного продукування мікробної маси змінюються періодами його затухання. Пояснити ці зміни коливаннями температури та вологості грунту не завжди вдається; можливо, в основі цього феномену лежать чисто популяційні причини.
Хоч щорічна продукція мікроорганізмів дуже висока, їх жива біомаса на конкретний момент значно менша. П.А.Кожевін та Д.Г.Звягінцев (1975) оцінюють її для дерново-підзолистих грунтів у 0,17 мг/г, сірих лісових – 0,28, чорноземів – 0,38 мг/г грунту. Маса мертвої органічної речовини мікроорганізмів завжди перевищує живу, і її роль в інтегральному продуційному процесі геосистеми вважається вельми істотною – більшість дослідників процесу гумусоутворення вважають, що мертві клітини мікроорганізмів є важливим матеріалом, з якого синтезується гумус.
Відтворення гумусу – не менш важлива ланка продуційного процесу в геосистемі, ніж продукування біомаси. Основним джерелом його формування є рослинний опад, екскременти тварин та клітини мікроорганізмів.
Швидкість та характер гуміфікації залежать від багатьох ландшафтно-екологічних факторів. З них найбільше значення мають кількість та склад рослинних решток, режим вологості та аерації грунту, його кислотність, видовий склад мікроорганізмів та інтенсивність їх діяльності, мінералогічний та механічний склад грунту тощо. Багатофакторність процесу гумусонагромадження зумовлює складність теоретичних математичних моделей цього процесу. Проте емпіричні та емпірико-теоретичні моделі були запропоновані. Так, Ф.М.Лісецький (1990), узагальнивши наявні дані щодо темпів гумусонагромадження в різних типах грунтів, вивів досить простий вираз, що описує формування гумусового горизонту грунтів – НГ (мм) на породах суглинкового механічного складу:
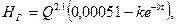
де Q – енергетичні витрати на ґрунтоутворення, Мдж/м2 за рік;
k –коефіцієнт типу грунту (у підзолистих грунтах k становить 0,00035, у чорноземах типових і звичайних – 0,00039, у чорноземах південних і темно-каштанових грунтів – 0,00044, у світло-каштанових та чорноземах карбонатних – 0,0004);
b – параметр інтенсивності формування гумусового горизонту в різні періоди формування грунту (у чорноземах і каштанових грунтах b становить 0,00034; у дерново-підзолистих та глейових грунтах – 0,00095);
t – час, роки.
Процес гуміфікації йде повільно (табл. 2) , причому з вихідної маси рослинного опаду на гумус перетворюється не більш як 10-30%.
Таблиця 2 (7) [1]. Середня швидкість формування гумусного горизонту грунтів, т/га (за Ф.М.Лісецьким, 1990)
| Грунти | Ступінь змитості грунту | ||
| слабка | середня | Сильна | |
| Дерново-підзолисті Чорноземи типові Чорноземи звичайні Чорноземи південні, темно-каштанові Світло-каштанові, чорноземи карбонатні | 0,3 0,5 0,4 0,4 0,2 | - 1,4 1,2 0,8 0,6 | - 2,6 2,0 1,8 0,8 |
Дата добавления: 2015-05-16; просмотров: 1005;